
© 2021 Gayle Morris Sweetland Center for Writing
Permission is required to reproduce material from this title in other
publications, coursepacks, electronic products, and other media.
Please send permission requests to:
Sweetland Center for Writing
1310 North Quad
105 S. State Street
Ann Arbor, MI 48109-1285
Excellence
in
Upper-Level Writing
2020/2021
e Gayle Morris
Sweetland Center for Writing
4
Table of Contents
Excellence in Upper-Level Writing
Winners list
Nominees list
Introduction
Prize for Excellence in Upper-Level Writing
(Sciences)
A Review of Titin: e Titans of Human Muscle
TGF-β1: Unraveling the Applications of a Versatile Cytokine
Prize for Excellence in Upper-Level Writing
(Social Sciences)
Research Proposal: Reevaluating the Economic Imperative to Learn
e Social Brain Hypothesis: An Evolutionary Explanation for Our
Big Brains
Prize for Excellence in Upper-Level Writing
(Humanities)
A Review of PLA’s Sustainability as the Future of Bioplastics
A Handful of Walnuts
6
7
9
11
28
40
60
74
89
5
Excellence in Upper-Level Writing 2020/2021
Sweetland Writing Prize Chair
Gina Brandolino
Sweetland Writing Prize Judges
Administrative Support
Andrew Bernard
Louis Cicciarelli
Jennifer Cummings
Domenic DeSocio
Marisol Fila
Sofya Khagi
Jie (Jackie) Li
Michael Martin
Wilson Merrell
Katy Rossing
Marissa Spada
eresa Tinkle
Field Watts
Laura Schulyer
Aaron Valdez
6
Winners List
Excellence in Upper-Level Writing (Sciences)
Puneet Dhatt
“A Review of Titin: e Titans of Human Muscle”
Nominated by Nicholas Garza, CHEM 353: Introduction to Biochemical Research
Techniques and Scientic Writing
Kateryna Karpo
“TGF-β1: Unraveling the Applications of a Versatile Cytokine”
Nominated by Nicholas Garza, CHEM 353: Introduction to Biochemical Research
Techniques and Scientic Writing
Excellence in Upper-Level Writing (Social Sciences)
Angelina Little
“Research Proposal: Reevaluating the Economic Imperative to Learn”
Nominated by Nancy Burns and Ben Goehring (GSI), POLSCI 381: Political
Science Research Design
Sahil Tolia
“e Social Brain Hypothesis: An Evolutionary Explanation for Our Big Brains”
Nominated by Andrew Bernard, ANTHRBIO 368: Primate Social Behavior
Excellence in Upper-Level Writing (Humanities)
Leah Marks
“A Review of PLA’s Sustainability as the Future of Bioplastics”
Nominated by Jimmy Brancho, WRITING 400: Advanced Rhetoric and Research,
Writing and Research in the Sciences
Julia Van Goor
“A Handful of Walnuts”
Nominated by Jamien Delp, ENG 325: Art of the Essay
7
Nominees List
Student
Dina Ahmad, Lauren Guido, &
Jaya yagarajan (collaboration)
Allegra Baird
Mikayla Lilly Bosma
Benjamin Bronkema-Bekker
Isabel Brush-Mindell
Zoe Bultman
Isabella Buzynski
Jay Campbell
Olivia Chang
Deeya Chaturvedi
Elizabeth Cho
Claire Pajka
Conor Raerty
Liam Corriston
Danielle Crasiuc
Hannah Davies
Julia Dean
Puneet Dhatt
Viviana Flores
Carter Fox
Dequan Gambrell
Catherine Garton
Tanner Gritzmaker
Kamryn Hayes
Alice Hill
Alice Hill
Vasili Ioannidis
Emily Johnson
Instructor
Michelle Bellino
Madelyn Cook
Marlon James Sales
Vedran Catovic
Emilia Askari
Rolf Bouma
Cameron Cross
Jennifer Metsker
Kevin Miller
Omolade Adunbi
Brian Remlinger
Julie Halpert
Julie Halpert
Gabriel VanLoozen
Sara Ahbel-Rappe
Andrew Bernard
Wilson Merrell
Nicholas Garza
Emily P. Lawsin, (Paulina Fraser, GSI)
Roy Clarke
Cameron Cross
Stephanie Preston
Christine Chalifoux
Rolf Bouma
Lynn Carpenter
Julian Schultz
Yehia Mekawi
Yehia Mekawi
8
Student
Kateryna Karpo
Daniella Kay
Alyssa Klotz
Alex Kocheril
Alexia LaLonde
Angelina Little
Gina Liu
Anthony Lohmeier
Christine Chia-Rong Lu
Anshuman Madhukar
Allison Malkowski
Brianna Marble
Leah Marks
Michael Moynihan
Celene Philip
Claire Ramsey
Sayan Raychaudhuri
Robin Rong
Samuel Rosenblum
Aryanna Rudolph
Jared Stolove
Sahil Tolia
Aria Trager
Julia Van Goor
Erin Walski
Levana Wang
Grin Zdrojewski
Keri Zhang
Michael Zhang
Instructor
Nicholas Garza
David Gold
Vedran Catovic
Omolade Adunbi
Allie Hirsch
Nancy Burns (Ben Goehring, GSI)
Omolade Adunbi
Omolade Adunbi
Jimmy Brancho
San Duanmu
Ben Hansen
Lynn Carpenter
Jimmy Brancho
Lisa Koo
Brian Remlinger
Lisa Koo
Roy Clarke
Emilia Askari
David Gold
Benjamin Hsu
Ashley Craig
Andrew Bernard
Omolade Adunbi
Jaimien Delp
Gabriel VanLoozen
Jaimien Delp
Wilson Merrell
San Duanmu
Julian Schultz
9
Introduction
All LSA undergraduates take at least one upper-level writing-intensive
course, often in their major or minor, and frequently in another eld of interest.
ose of us who teach these courses discover anew each term how a focus on
writing improves students’ thinking, learning, and rhetorical creativity. At the end
of each term, we applaud our students’ various achievements and the ways they
have grown by applying themselves seriously to the hard work of designing a study,
conducting research, addressing an audience eectively, crafting a multimedia
essay, rening their prose style, reading and giving feedback on peers’ drafts,
or engaging with counter-arguments. ese are rewarding courses to teach, as
witnessed by the signicant number of faculty from all disciplines who participate
robustly in the upper-level writing program each year.
Each year, faculty and graduate student instructors encourage
undergraduates to submit their very best essay for the Upper-Level Writing Prizes.
Fellows in the interdisciplinary Sweetland Seminar for Writing Pedagogy read
the submissions and rank them according to their overall excellence. is is an
intellectually interesting exercise and generates considerable discussion about
what we value when we read students’ work. e Fellows this year described for
each other the qualities they admired in the submissions. Here are some of the
ways they dened excellent writing: the relationship between existing scholarship
and a student’s position is clearly detailed and persuasive; the argument is
complex without loss of clarity or purpose; the piece presents a novel argument or
assertion (e.g., coming to their own conclusions based on multiple texts); quotes
are thoughtfully introduced and integrated into the argument; the evidence
is compelling; and the writers use rich, evocative language. All of the essays
submitted were outstanding, and all of the students should feel very proud of
what they accomplished.
10
is volume showcases the prize-winning essays, which are truly
impressive. ey witness to the robust intellectual life of the university, and to
the splendid courses and instructors who inspired and supported the writers’
accomplishments. Each essay speaks to how much our students contribute to the
creation of new knowledge.
anks are due to the many people who made this volume possible.
e Senior Fellows who thoughtfully judged the essays are Louis Cicciarelli,
Sweetland Center for Writing; Jennifer Cummings, Psychology, Biopsychology;
Sofya Khagi, Slavic Language and Literatures; and Jie (Jackie) Li, Earth and
Environmental Sciences. e Junior Fellows (Graduate Students) are Andrew
Bernard, Anthropology; Domenic DeSocio, Germanic Language and Literature;
Marisol Fila, Romance Languages; Michael Martin, Slavic Language and
Literatures; Wilson Merrell, Psychology; Katy Rossing, English Language and
Literature; Marissa Spada, Film, Television, Media; and Field Watts, Chemistry.
Much gratitude is also due to Aaron Valdez, who designed this volume; Laura
Schuyler, who coordinated the submission and judging process; and Gina
Brandolino, who chaired the Sweetland Prize Committee and edited the volume.
Finally, thank you to the students and instructors who strive for—and achieve—
excellence in writing and writing pedagogy.
eresa Tinkle
Sweetland Center for Writing
Arthur F. urnau Professor and Professor of English
11
Excellence in Upper-Level Writing
(Sciences)
A Review of Titin: e Titans of Human Muscle
by Puneet Dhatt
From CHEM 353: Introduction to Biochemical Research Techniques
and Scientic Writing
Nominated by Nicolas Garza
In his review paper, Puneet struck an elegant balance between the scientic
format and narrating the results. Many scientists struggle to make their written
works ow naturally while still conveying the content, and Puneet excelled at that
in his exploration of the protein titin. His introduction and function sections
were particularly engaging, and at times I forgot I was reading a student paper
instead of a published article.
-- Nicolas Garza
12
A Review of Titin: e Titans of Human Muscle
Abstract:
Titin is the largest protein known to exist in the human body. Composed of
more than 24,000 amino acids, titin has a remarkable molecular weight of over
33,000kDa. As such a large protein, it would be logical to assume titin has many
dierent subdomains. However, having recently fully discovered the structure
of titin, we now know that 90% of these domains are Ig and Ig-FNIII. Titin’s
main function is structural, providing a passive force with muscle contraction that
allows the myocytes’ sarcomeres their elasticity. Recently, many strides have been
made in the eld of titin’s structure and function. In this review, we analyze such
recent strides and how they convey the essential role titin plays in the body. en,
we will examine how titin’s structure can mutate and how these mutations lead to
many neuromuscular diseases.
Introduction:
Central to Greek mythology is the old god, the Titan, Atlas. A Titan
leader, Atlas is punished for his uprising against Zeus, condemned to hold up
the Earth. Allegorical to this mythic great, the protein titin exists in our muscles,
condemned by evolution to provide them with the elasticity they need to function.
It has been over 40 years since titin (also known as connectin) was discovered and
characterized.
1,2,3
After its discovery as a novel elastic element of myobrils,
1
it was
found that titin was the third principle element of the myobril, suggesting a new
three lament model instead of the previously accepted two lament model.
4,5
Titin falls into the class of structural proteins, and though it is in many cells, it
mainly functions as a component of the myocyte’s sarcomere.
4,20
Titin’s discovery
meant the characterization of the now largest known protein (~3MDa).
6
It has
been known since its discovery that titin is an important elastic element of striated
muscle assembly -known as the “muscular spring”.
7
Specically, titin’s >33000
residues can be found in the sarcomere where this giant protein spans half of the
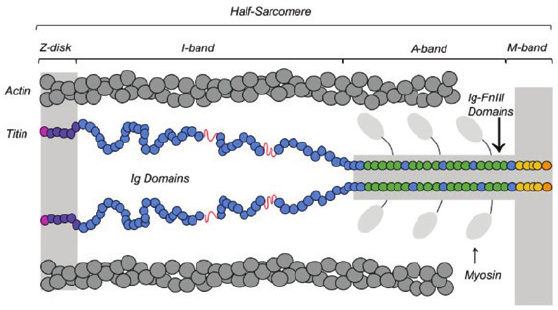
sarcomere from the Z-disk to the M-band, as seen in Figure 1.
8
Being such a large
13
protein, titin has many domains as it stretches across the half-sarcomere, each
with its own unique function as will be described (Figure 1).
Many investigations into this protein’s structure and function have begun
to elucidate how titin’s cycling works and contributes to the contraction and
relaxation of the sarcomere.
7
Starting with structure, this review will examine the
role of titin’s immunoglobulin and bronectin domains and their contribution
to the muscle contraction mechanism.
6,7,9
en, we will move on to examine
advances in the eld studying titin’s function as a large-scale regulatory node for
muscle cells remodeling and signaling pathways.
10,11
In addition, we will discuss
di erent mutations and titin variants that can lead to the emergence of disease,
especially in terms of titin’s developing role in causing neuromuscular disorders.
8
Finally, titin’s newfound role in contributing to heart disease and failure will be
focused on.
12,13,14
As heart disease is the leading cause of death in the United States
today, this eld could yield promising information for future therapies. In order
for this to occur, rst the true mechanism of how titin causes these diseases has to
be elucidated, for which there is currently little information.
13
Figure 1: e titin lament stretches across the half-sarcomere. Titin stretches
from the Z-disk (purple), where it is anchored by its N-terminus (pink). e
lament extends to its C-terminus (orange), where it is anchored (yellow) to the
M-band. e titin lament is made up of folded Ig and Ig-FNIII domains (blue
14
and green respectively) and intrinsically disordered structures (N2-B and PEVK,
in red). us, titin has 4 regions: Z-disk (purple), tandem Ig domains (blue),
A-band region (green and blue), and the M-band (yellow).
20
Titin Structure and Subdomains
Spanning the half-sarcomere, titin is the largest known protein. Most of
this protein, about 90%, is composed of Ig and bronectin (Ig-FNIII) domains.
6
Conventionally, this gigantic protein is understood in smaller groupings of
domains known as regions.
18,19
ere are 4 such regions: the Z-disk region, the
M-band region, the I-band region, and the A-band region, organized by region
and function in the sarcomere (Figure 1).
19
In the Z-disk domain, the titin
lament is anchored to the Z-disk, allowing it its stability and ability to provide
passive tension with the muscle contraction driven by the active myosin and actin
lament (Figure 1).
19
e Z-disk is also the location of the N-terminus of the
protein (Figure 1).
19
e M-band region encompasses another titin anchoring
domain and its C-terminus (Figure 1).
19
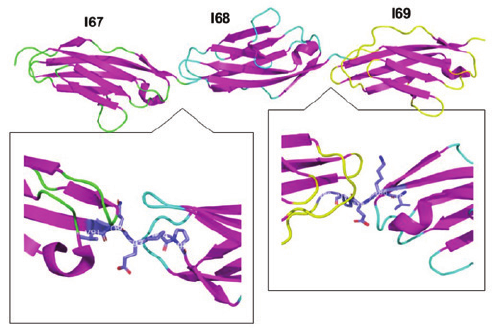
Figure 2: Structure of Ig domains. Figure created in Pymol. I67-I69 are
shown here from the I-band elastic region of the titin lament. (Left) the
I67(green)-I68(cyan)linker domain is the VQEPP motif (purple). (Right) the
I68(cyan)-I69(yellow)linker domain is the VKEPP motif (purple). NOTE:
15
image is positioned such that I69(yellow) is on the left for better visualization
of the linker bonds (pdb: 2RIK).
e most important regions to titin’s function are the I-and A-band
regions. e I-band region is composed of many dierentially spliced Ig
(immunoglobulin) domains in repetition and is responsible for the elasticity of
titin (Figure 2).
19,20,25
is means that there are a varying number of Ig domains
that make up this region. Specically, this number is thought to vary between
myocytes of dierent muscle groups, as well as between individuals.
20
Principally,
this region confers titin its elasticity, a dening characteristic of the lament we
call the “muscular spring”.
20
e orientation of these domains, when the muscle
is relaxed, is similar to that seen in Figure 2, with the Ig domains curled up and
free to move around.
19
e exact mechanism for the muscle contraction rebound
will be discussed later on. Understanding titin’s role as the elastic element in the
sarcomere is essential to comprehending the protein’s function itself. In addition
to these Ig domains,there are also 2 intrinsically disordered domains found in the
I-band region, N2-A/B and PEVK.
19,20
ese regions allow an additional spring-
like nature to the protein.
19
e N2-domain comes in 3 avors, N2-A, N2-B, and
N2-BA (most commonly found in the heart).
8
Figure 2 shows the binding between
these Ig domains in the I-band.
25
ere is a dedicated linker domain that varies for
dierent Ig domains, but that is used to hold the domains together(Figure 2). In
the I67-69 stretch, these motifs consist of VQEPP and VKEPP, or a hydrophobic
sandwiching of hydrophilic residues (Figure 2).
25

16
Figure 3: e protein structure and of Fibronectin domains. (A) Figure created
in Pymol. Representation of Fibronectin domain of A77, along with linker
domain S100-E101-R102 (green).
16
(B) Domains A77 (left) and A78 (right)
are connected by a three-residue hydrophilic linker S100-E101-R102 on A77
connecting to H128, S132, K182, D185 on A78.
16
is forms a rich network of
polar interactions that holds these two domains rigidly anti-oriented with respect
to each other (pdb:3LPW).
e A-band region is mostly composed of Ig-FNIII ( bronectin type III)
domains.
19
e main role of this region of the lament is to tightly bind the thick
lament (myosin) to allow for muscle rebound.
19
How the Ig-FNIII domains link
to each other is visualized in Figure 3. In Figure 3, the linker domain is seen to be
a hydrophilic S100-E101-R102 motif.
16
is motif of A77 associated with more
hydrophilic residues on A78 (such as H128 and S132).
16
Such a rich network of
polar associations di ers from how Ig domains link to each other, as they use both
hydrophobic and hydrophilic interactions. is increase in hydrophilic association
allows the Ig-FNIII domains to remain more rigid, and conversely the Ig domains
of the I-band to remain more uid or elastic.
16
is rigidity allows the domains to
17
remain anti-oriented with respect to each other, an element of their rigid structure
that is essential to the function of the A-band of the lament(Figure 3).
16
e main method by which we know these structures of titin is
through x-ray crystallographic studies.
25
Both Figure 2 and 3 were derived from
such studies. e other main method of structural analysis is through NMR
studies, including 2D NMR studies such as TOCSY and NOESY.
6
e 2D
NMR studies in particular reveal through space (NOESY) and through bond
(TOCSY) interactions that are central to elucidating interacting pairs of amino
acid side chains that serve key roles in the tertiary and quaternary structure of the
polypeptide sequence.
Titin Function and ree-Filament Muscle Contraction Mechanism
As previously mentioned, titin confers elasticity to the sarcomere through
its I-band domains.
19,20
is elasticity is key to supply the sarcomere with passive
force that compliments the active contraction driven by actin and myosin.
7
Before
the discovery of titin, this passive force was contributed to structural elements of
the sarcomere such as collagen
.7
e discovery of titin also revolutionized thinking
about the structure of the sarcomere.
7
Initially, the two-lament model of muscle
contraction was accepted.
4,5
In this two-lament model that is commonplace in
an introductory course in biology, actin is pulled by myosin to bring together
the Z-disk and M-line, a process that requires metabolic input to power (Figure
1). However, after titin’s discovery, a three-lament model–one that depends on
actin, myosin, and titin – has become increasingly popular.
5
e mechanism of
titin’s inuence on muscle contraction has not yet been entirely realized but there
have been some major advancements in this eld recently thanks to atomic force
microscopy (AFM) studies.
18,23
ese studies use magnetic tweezers for single
molecule microscopy that allow nanoscale mechanics of titin to be realized.
18,23
ough not entirely realized, we do now understand the basics of titin’s
role in muscle contraction. In its relaxed state, titin exists stretched out, with
its Ig domains elongated due to the sarcomere pulling against titin to stretch

18
the muscle to its relaxed state. is passive tension that is supplied by titin is
the “passive force” that is often referenced in titin’s function.
7
As the muscle
contracts, cross bridges are formed between actin and myosin laments and the
force on titin decreases.
18,19,22
e force decreasing as the sarcomere contracts
allows titin’s I-band Ig domains to fold.
22
is folding is favorable for the domain
but was disallowed by the stretched conformation of the relaxed muscle.
18,19,22
Furthermore, this folding is cooperative, such that one domain spontaneously
folding will increase the probability that those around it will also fold.
22
Figure 4: Entropic Recoil of Titin. When there is a force to maintain stretched,
during muscle relaxation, titin remains stretched out (left). However, as the
muscle contracts, the lament coils, and as a result has many more conformations
open to it that it can sweep through (right). is increase in conformations to
sweep through means a favorable increase in the entropy of the system.
Another mechanistic reasoning for titin contraction during muscle
contraction is that as the sarcomere shrinks in size due to the active laments
work, there is an entropic driving force for titin to rebound.
6,18
In its relaxed state,
sarcomeric titin is disallowed from occupying many microstates, as the stretched
conformation of the lament is more controlled and rigid. is low microstate
count means a lower entropy for this state. As the sarcomere contracts, titin is
no longer forced to hold such a stretched conformation. As such, many more
microstates open up, increasing the entropy of the lament. is can be seen in
Figure 4 wherein the straight conformation on the left has a single conformation
but there are many more conformations that open up as the lament coils. is
coined “entropic rebound” has been used to explain titin passive force mechanics
as well.
6,18
In reality, though both of these mechanisms provide some insight

19
into titin’s rebound reasoning. rough AFM studies we know that the folding
contraction of just one Ig domain generates twice the work of just the entropic
recoiling in the physiological 4-15pN force range wherein titin operates.
18
is
means that current evidence points toward the folding work of the individual Ig
domains driving titin rebound.
8
Titin’s Role as a Regulatory Node
Figure 5: Cardiomyocyte signaling pathways involving titin. Cardiac forms
of titin (N2-BA and N2-B) are here seen as binding partners and involved in
hypertrophic signaling pathways. Keywords: ERK2, extracellular signal-regulated
kinase 2; G, small G-protein; MLP, muscle LIM protein; MURF1/2, muscle
20
RING nger protein-1/2; P, titin phosphorylation site; sGC, soluble guanylyl
cyclase. (Figure from Krüger and Linke, 2011).
10,21
Along with its key structural features, titin has a newly discovered and
emerging role as a key regulatory node.
10,21
Seen in Figure 5, many signaling cascades
end on the titin lament, especially in cardiomyocyte signaling.
10
ese cascades
are initiated by many peptides landing on dierent cellular receptors. Recently,
titin’s role in cellular signaling cascades in cardiomyocytes has been elucidated.
10
During fetal development, it has been discovered that the N2BA/N2B ratio
increases, increasing the stiness of titin in the heart, and therefore the passive
force for contraction.
10
In addition, titin phosphorylation and its modulations
have recently been uncovered. Phosphorylation is a common way to regulate
protein function. e addition or removal of a phosphate group leads to changes
in a protein’s tertiary or quaternary structure such that its function is modulated.
As such, phosphorylation is key to titin’s function and regulation.
10,21
In fact, it is
phosphorylation of the XSPXR motif of titin that initiates myogenesis.
10
e titin
lament needs this phosphorylation to recruit the necessary cofactors to initiate
the construction of myobrils.
10
Additionally, not only serving as a scaold, titin
also organizes the sarcomere.
10
At titin’s carboxyl terminus in the Z-disk, there
is a Mis-4 insertion sequence.
10
is sequence contains 4 KSP motifs that are
phosphorylated to regulate SH3 (Src homology 3) binding for Bin1, a protein
that organizes the sarcomere.
10
Phosphorylation of titin can also control its passive mechanistic qualities.
For example, phosphorylation of S469 in the N2-B intrinsically disordered
structure can reduce titin’s passive tension by up to 20%.
10,21
Furthermore,
phosphorylation can also be used to increase passive tension in cardiomyocytes.
10,21
is occurs through PKCα phosphorylation of COOH domain of the PEVK
structure of titin.
10,21
ese transient phosphorylation and dephosphorylation
cascades can modulate titin in a beat-by-beat manner in response to extracellular
signals.
10
Not only does titin get phosphorylated to be regulated but titin does
21
have a kinase domain, the titin kinase domain (TK).
10
Phosphorylation of a
tyrosine residue in this domain initiates a complex activation process. TK is an
autoinhibited S/T kinase.
10
e exact mechanism of its activation is unknown,
as whether the active site tyrosine is activated through auto-phosphorylation
or phosphorylated by some coactivatoris unknown.
10
is TK domain has also
been suggested to participate in phosphorylation of Z1/Z2-BP telethonin,
found at titin’s N-terminus.
10
However, although there is data to suggest this
phosphorylation is mechanical stress adaptations, its full eect is not yet known.
10
Apart from phosphorylation, titin also plays a key role in muscle
hypertrophic gene regulation.
10,11
Hypertrophy is when an organ is enlarged due
to increases in cell size. Titin not only is a large protein but has a large network
of protein it associates with that link titin signaling to pathways regulating
hypertrophy.
10
TitinZ1/Z2 (Z-disk region) domains tightly bind a telethonin/T1
cap. Such an interaction links titin to the muscle LIM protein, which has many
locations in the myocyte including the nucleus wherein it acts as a coactivator
of transcription.
11
ere are also negative regulators of gene regulation found in
the signaling of titin through the node on the I-band in the N2-A region.
10,11
Binding proteins bind to the I80/81 region.
10,11
ese are MARPs (muscle ankyrin
repeat proteins).
10,11
ey move to the nucleus in response to mechanical strain
and cause negative gene regulation.
10,11
Finally, in the M-band, A168-170 interact
with MURF1 and 2. MURF1/2 can translocate to the nucleus in response to stress
signals and mediate transcription as well as seen in Figure 5.
10,11
All of this reveals
titin’s role as a regulatory crossroads for signaling, especially in cardiomyocytes.
10,11
Titin Mutations and Associated Diseases:
e polypeptide titin is encoded by the TTN gene.
8
is gene is very large,
containing 363 exons.
8
Mutations in the TTN gene have already been associated
with many cardiomyopathies.
8
Such a discovery tracks with titin’s essential role
in cardiomyocyte signaling as described previously. ese titin-associated diseases
include: Late-onset autosomal dominant tibial muscular dystrophy (TMD),

22
Young or early adult onset recessive distal titinopathy, Congenital centronuclear
myopathy (CNM), Early-onset myopathy with fatal cardiomyopathy (EOMFC),
Multi-minicore disease with heart disease (MmDHD), Childhood-juvenile
onset Emery-Dreifuss-like phenotype, and adult onset recessive proximal
muscular dystrophy.
8,24
All of these diseases associated with titin mutations lead
to phenotypes of muscular degeneration or dysfunction, revealing titin’s key
role in muscle contraction and the sarcomere’s structure. Speci cally, we have
correlated numerous diseases involving fetal and adult-onset heart failure with
titin mutations. As heart failure is the leading cause of death in the United States
today, titin could play a pivotal role in our understanding and development of
treatments for these illnesses.
Figure 6: Disease states correlated to their relative mutations on each allele of
titin. Small missense mutations can dramatically change the titin structure such
that many di erent kinds of muscular disorders develop.
8
Before recently, with the advancements in next-generation sequencing
techniques (NGS), it was not possible to sequence the entire TTN gene.
24
However,
even though it is now possible these data can be hard to interpret, as many genetic
23
variants of TTN do not have a known correlated disease phenotype.
8
Figure 6
shows the known mutation-disease pairings along with where and what time of
mutation occurs.
8
In such tables, we see that titin mutations can be grouped into
TMD, titinopathic, and LGMD categorizations. ese categorizations are made
based on disease phenotypes, but as seen in Figure 6 are also correlated through
their respective mutation sites. For example, mutations in the M10 domain lead
to phenotypes associated with TMD (Figure 6).
Many of these diseases were of unknown cause until these links to the
TTN gene were established.
8
ere are many key outcomes to such ndings.
Principal among these is the stressing of the essentiality of titin.
15
Titin is a key
element of the sarcomere, and as such is central to the function of the heart.
15
It
is these disease correlations begin to show us just how essential a role titin plays in
our bodies. We also begin to learn titin’s inuence outside of just being a structural
scaold for the sarcomere but extending to its roles in signaling pathways.
Aside from NGS that allows us to nally eectively sequence the entire
TTN gene, other techniques have also revolutionized our understanding of titin:
iPS-CM and CRISPR studies. iPS-CM stands for induced pluripotent stem cell-
derived cardiomyocytes. In this technique, disease patients’ cells are taken, and
pluripotent stem cells are derived from their DNA. us, a line of diseased cells
is sequenced and studied for morphological as well as functional deciencies
that underlie the diseased phenotype. With advances in CRISPR technology,
scientists have learned how to forego the need for a diseased patient and now
can target certain parts of the TTN gene to upregulate and downregulate its
specic transcription.
17
As such, now we no longer have to wait for patients with a
disease variant and can instead induce our own iPS cells with many more specic
variations in the TTN gene. is allows for more testing and more specic analysis
in which mutations in which position along the gene result in which phenotype.
ere is still much work to be done in this eld. Much is unknown about
these disease’s mechanism, including central questions like how or why they occur.
However, much of the foundations have recently been lain and are propelling this
24
eld quickly into the future. is includes developments and utilization of new
cutting-edge techniques as well as an initial foray correlating TTN mutants with
disease phenotypes.
Conclusion
Less than a half century ago we did not know titin existed. Now, we
have discovered it mainly consists of Ig and Ig-FNIII domains, and its role as
an elastic structural protein that provides the scaold upon which the sarcomere
is constructed. Titin has four denitional regions: Z-disk, I-band, A-band, and
M-band regions. Titin is intricately and necessarily linked to the mechanism of
muscle contraction, providing passive tension for the sarcomere that with the
active sliding lament elements of actin and myosin allow for muscle contraction.
e drive towards contraction is provided by the favorable folding and entropic
rebound associated with Ig domains in the I-band region, rather than metabolic
input like myosin. anks to recent advancements that have led to NGS and
the CRISPRa/i system, we also have begun to understand how mutations in the
titin gene, TTN, can cause disease. Titin so far has been particular associated
with neuromuscular diseases such as muscular dystrophy. Finally, titin has been
indicated as a key regulatory node in myocytes. From phosphorylation cascades
that modulate titin to those in which titin regulates other proteins, titin has been
seen to be a key node in protein regulation. However, titin is also key in genetic
regulation with its interactions with MURF and LIM proteins.
Future studies of titin hold a lot of promise. For example, work to
understand underlying titin-associated disease mechanisms could hold the key
to unlocking a myriad of treatments for muscular and cardiac disorders that
currently have none. Such studies could include exploration of the sarcomere’s
integrity due to titin mutations or force microscopy studies focusing on the eect
of titin mutations on titin’s ability to provide passive tension to the sarcomere.
Moreover, further sequence testing of muscular disorders looking for titin mutants
could provide integral information to treating a host of diseases that currently
25
are idiopathic as well as reveal clinical markers to identify these illnesses much
earlier. Finally, future studies could help us understand the evolutionary ladder of
life on Earth as elucidating the connection between titin and titin-like proteins
found in dierent mammalian species could be used as an evolutionary marker.
ese studies could also reveal conserved domains across species and time that
would provide key details to which domains of titin are the most essential for its
function.
References:
1. Maruyama, K.; Matsubara, S.; Natori, R.; Nonomura, Y.; Kimura, S.,
Connectin, an elastic protein of muscle. Characterization and Function. J
Biochem 1977, 82, 317-37.
2. Wang, K.; McClure, J.; Tu, A., Titin: major myobrillar components of
striated muscle. Proceedings of the National Academy of Sciences of the United
States of America 1979, 76, 3698-3702.
3. Maruyama, K.; Kimura, S.; Ohashi, K.; Kuwano, Y., Connectin, an elastic
protein of muscle. Identication of “titin” with connectin. J Biochem 1981,
89, 701-9.
4. Magid, A.; Law, D. J., Myobrils bear most of the resting tension in frog
skeletal muscle. Science 1985, 230, 1280-2.
5. Huxley, H.; Hanson, J., Changes in the cross-striations of muscle during
contraction and stretch and their structural interpretation. Nature 1954,
173, 973-6.
6. Goll, C.; Pastore, A.; Nilges, M. e three-dimensional structure of a type I
module from titin: a prototype of intracellular bronectin type III domains.
Structure, 1998, 6, 1291-1302.
7. Herzog, W., e multiple roles of titin in muscle contraction and force
production. Biophysical Reviews 2018, 10, 1187-1199.
26
8. Savarese, M.; Sarparanta, J.; Vihola, A.; Udd, B.; Hackman, P. Increasing
Role of Titin Mutations in Neuromuscular Disorders. Journal of
Neuromuscular Diseases, 2016,3, 293-308.9) Machado C.; Sunkel C.E.;
Andrew D. Human autoantibodies reveal titin as a chromosomal protein. J.
Cell Biol. 1998, 141, 321-333.
9. Krüger, M.; Linke, W. e Giant Protein Titin: A Regulatory Node at
Integrates Myocyte Signaling Pathways. Journal of Biological Chemistry,
2011, 286, 9905-9912.
10. Krüger, M.;Kötter, S., Titin, a Central Mediator for Hypertrophic Signaling,
Exercise-Induced Mechanosignaling and Skeletal Muscle Remodeling.
Frontiers in Physiology, 2016, 7, 1-8.
11. Hinson, J. T.; Chopra, A.; Nassi, N.; Polacheck, W. J.; Benson, C. C.;
Swist, S.; Gorham, J.; Yang, L.; Schafer, S.; Sheng, C. C.; Haghighi, A.;
Homsy, J.; Hubner, N.; Church, G.; Cook, S. A.; Linke, W. A.; Chen, C.
S.; Seidman, J. G.; Seidman, C. E., HEART DISEASE. Titin mutations in
iPS cells dene sarcomere insuciency as a cause of dilated cardiomyopathy.
Science 2015, 349, 982-6.
12. Kellermayer, D.; Smith, J. E., 3rd; Granzier, H., Titin mutations and muscle
disease. Pugers Arch 2019, 471, 673-682.
13. Zile, M. R.; Baicu, C. F.; Ikonomidis, J. S.; Stroud, R. E.; Nietert, P.
J.; Bradshaw, A. D.; Slater, R.; Palmer, B. M.; Van Buren, P.; Meyer,
M.; Redeld, M. M.; Bull, D. A.; Granzier, H. L.; LeWinter, M. M.,
Myocardial stiness in patients with heart failure and a preserved ejection
fraction: contributions of collagen and titin. Circulation 2015, 131, 1247-
59.
14. Azad, A.; Poloni, G.; Sontayananon, N.; Jiang, H.; Gehmlich, K., e giant
titin: how to evaluate its role in cardiomyopathies. Journal of muscle research
and cell motility 2019, 40(2), 159-167.
15. Bucher, R. M.; Svergun, D. I.; Muhle-Goll, C.; Mayans, O., e structure
of the FnIIITandem A77-A78 points to a periodically conserved architecture
in the myosin-binding region of titin. J Mol Biol 2010, 401(5), 843-53.
16. Karakikes, I.; Ameen, M.; Termglinchan, V.; Wu, J. C., Human induced
pluripotent stem cell-derived cardiomyocytes: insights into molecular,
cellular, and functional phenotypes. Circulation research 2015,117(1),
80-88.
27
17. Eckels, E. C.; Haldar, S.; Tapia-Rojo, R.; Rivas-Pardo, J. A.; Fernández,
J. M., e Mechanical Power of Titin Folding. Cell Reports 2019, 27(6),
1836-1847.e4.
18. Eckels, E. C.; Tapia-Rojo, R.; Rivas-Pardo, J. A.; Fernández, J. M., e
Work of Titin Protein Folding as a Major Driver in Muscle Contraction.
Annual review of physiology 2018, 80, 327-351.
19. Freundt, J. K.; Linke, W. A., Titin as a force-generating muscle protein
under regulatory control. Journal of Applied Physiology 2018, 126(5), 1474-
1482.
20. Koser, F.; Loescher, C.; Linke, W. A., Posttranslational modications of titin
from cardiac muscle: how, where, and what for? e FEBS Journal 2019,
286(12), 2240-2260.
21. Mártonfalvi, Z.; Bianco, P.; Naftz, K.; Ferenczy, G. G.; Kellermayer, M.,
Force generation by titin folding. Protein science: A Publication of the Protein
Society 2017, 26(7), 1380-1390.
22. Rief, M.; Gautel, M.; Gaub, H. E., Unfolding forces of titin and bronectin
domains directly measured by AFM. Adv Exp Med Biol 2000, 481, 129-36;
discussion 137-41.
23. Savarese, M.; Maggi, L.; Vihola, A.; Jonson, P. H.; Tasca, G.; Ruggiero, L.;
Bello, L.; Magri, F.; Giugliano, T.; Torella, A.; Evilä, A.; Di Fruscio, G.;
Vanakker, O.; Gibertini, S.; Vercelli, L.; Ruggieri, A.; Antozzi, C.; Luque,
H.; Janssens, S.; Pasanisi, M. B.; Fiorillo, C.; Raimondi, M.; Ergoli, M.;
Politano, L.; Bruno, C.; Rubegni, A.; Pane, M.; Santorelli, F. M.; Minetti,
C.; Angelini, C.; De Bleecker, J.; Moggio, M.; Mongini, T.; Comi, G. P.;
Santoro, L.; Mercuri, E.; Pegoraro, E.; Mora, M.; Hackman, P.; Udd, B.;
Nigro, V., Interpreting Genetic Variants in Titin in Patients With Muscle
Disorders. JAMA neurology 2018,75(5), 557-565.
24. von Castelmur, E.; Marino, M.; Svergun, D. I.; Kreplak, L.; Ucurum-
Fotiadis, Z.; Konarev, P. V.; Urzhumtsev, A.; Labeit, D.; Labeit, S.; Mayans,
O., A regular pattern of Ig super-motifs denes segmental exibility as the
elastic mechanism of the titin chain. Proceedings of the National Academy of
Sciences of the United States of America 2008, 105(4), 1186-1191.
28
TGF-β1: Unraveling the Applications of a Versatile
Cytokine
by Kateryna Karpo
From CHEM 353: Introduction to Biochemical Research Techniques
and Scientic Writing
Nominated by Nicolas Garza
CHEM 353 student Kateryna Karpo did an excellent job in review of
the protein TGF-β. She did an outstanding job of creating and incorporating
gures into her paper. I as an instructor often nd students to struggle with gure
incorporation, but Kateryna excelled at this. Her signaling and disease sections
were concise yet detailed at the same time, which is dicult to achieve and an
indication of an excellent scientic writing piece.
-- Nicolas Garza
Excellence in Upper-Level Writing
(Sciences)
29
TGF-β1: Unraveling the Applications of a Versatile
Cytokine
Abstract
e TGF-β superfamily is composed of 35 members with a vast array of signaling
functions across both tissue homeostasis and cellular growth cycles. is review
highlights the most populous member of the TGF-β subfamily, TGF-β1. e
structure of this isoform contains both unique and conserved elements with
respect to its alternate isoforms, TGF-β2 and TGF-β3. Once synthesized,
TGF-β1 is released from the extracellular matrix and is ready to participate in
signaling pathways. e isoform begins by forming SMAD complexes, ultimately
localizing to the nucleus to behave as a transcription factor. e signaling patterns
of TGF-β1 are highly pleiotropic in nature, presenting dierent eects across
cellular environments and upon impact from alternate pathways. As these signaling
cascades are widespread across body systems, their disruption may result in the
onset of many human diseases, commonly causing the formation of both solid and
hematopoietic tumors. is opens the potential for TGF-β as a potential target
point for cancer therapy, with its inhibition slowing the uncontrolled cell growth.

30
Introduction
Transforming growth factor beta (TGF-β) is a multifunctional cytokine exhibiting
major roles in development and homeostasis of body tissues, as well as cellular
functions including proliferation, di erentiation, and apoptosis.
1
Figure 1. Diagram of the TGF-β superfamily members. (A) A schematic representation
of the TGF-β superfamily subfamilies is presented. TGF-β: transforming growth factor
beta; GDF: growth and di erentiation factor; ACT: activin; INH: inhibin; GDNF: glial-
derived neurotrophic factors; BMPs: bone morphogenetic proteins. (B) A web of the 35
TGF-β superfamily members organized by structural relationship is shown, placing the
TGF-β1 isoform (red) in the context of related growth factors and cytokines.
e TGF-β superfamily consists of ve main subfamilies – the activin/inhibin,
bone morphogenetic protein (BMP) & receptor, growth di erentiation factor
(GDF), glial cell line-derived neurotrophic factor (GDNF), and TGF-β families
(Figure 1A).
2
e members of these six subfamilies have varying evolutionary
relationships based on their structures, and this dictates which subfamily they
categorically fall under (Figure 1B). TGF-β was rst discovered while studying
the ability of polypeptide growth factors to implement autocrine secretion for the
induction of uncontrolled cell growth. When the sarcoma growth factor (SGF)
polypeptide was found in rat broblasts, studies indicated that SGF consists of two
substances – TGF-β and TGF-α.
1
Roberts and Sporn later characterized TGF-β
as a polypeptide secreted to promote the growth of broblasts and production
of collagen. After several years, it was found to inhibit cell proliferation, directly
connecting TGF-β inhibitors to the onset of cancer pathology.
1

31
e TGF-β1 isoform of the TGF-β subfamily is the most common of
three isoforms – TGF-β1, TGF-β2, and TGF-β3. While these isoforms share
signi cant structural features, they are synthesized in di erent organs for the
execution of varying tasks.
3
Speci cally, TGF-β1 is most often found in bone,
skin, and cartilage tissue with its key roles in di erentiation, while TGF-β2 is
expressed by neurons and is mainly responsible for the autonomic proliferation of
these cells. TGF-β3 is responsible for epithelial-mesenchymal interactions and is
most commonly found in lung and palate tissues. Disruptions in the functioning,
production, or signaling of TGF-β proteins, as well as interference with the
downstream products of TGF-β pathways, contribute to the development of
many diseases. is paper will investigate the structure and localization of the
TGF-β1 isoform; review the dominant TGF-β SMAD signaling pathway; cover
the TGF-β signaling disruptions which amount to cancerous uncontrolled cell
growth; and conclude by discussing TGF-β as a therapeutic target in cancer
treatment.
TGF-β1 Key Structural Features
e 35 members of the TGF-β
superfamily vary widely in structural
and evolutionary relationship (Figure
1B). Looking at the TGF-β subfamily,
the three existing isoforms share a
signi cant conserved component in
their sequences, however each has
ngerprint structures which makes
it unique. Speci cally, the precursor
polypeptide from which TGF-β1,
TGF-β2, and TGF-β3 are derived is initially composed of 300-600 amino acids
(Figure 2). is initial structure can be broken down to a variable N-terminus
fragment of approximately 200-500 amino acids, as well as a conserved C-terminus
fragment of about 110 amino acids.
4
Comparing the sequence similarity of the
Figure 2. Structure of proStructure of pro-
-TGFTGF--β1β1. e two beta--pleated
pleated sheets in the center highlight the
TGF-β knot, marking a distinct feature of
the TGF-β subfamily member structures.
PMID::21677751. PDB: 3RJR.
32
three isoforms, TGF-β1 and
TGF-β2 share about 71.4%
of their primary structures,
TGF-β1 and TGF-β3 share
about 76%, and TGF-β2 and
TGF-β3 share around 80%.
2
Looking at secondary and
tertiary structural features, the
entire TGF-β superfamily shares
seven cysteine residues that
ultimately form disul de bonds.
e following speci c structural
similarities are responsible for
de ning the subfamilies. For the
TGF-β subfamily, a conserved structure called the “TGF-β knot,” or a set of two
beta-pleated sheets anchored by a core of six cysteine residues, is seen across the
three members, with its seventh cysteine functioning to dimerize the monomers
through an interchain disul de bond.
4
is knot is visible in the center of the
largely unmodi ed dimeric pro-TGF-β1 structure (Figure 2, Figure 3B). e
structure of TGF-β1 in particular can be described as a homodimer of two 112
amino acid polypeptide chains linked through a disul de bond, amounting to a
total molecular weight of 25 kDa.
2
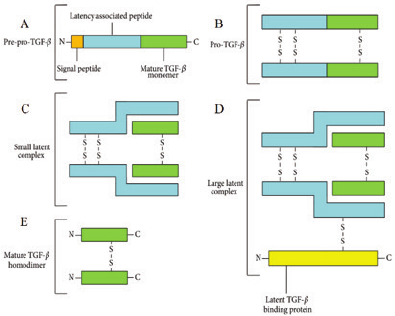
TGF-β1 Synthesis
ese structural features of TGF-β1, speci cally its ability to form disul de
bonds, directly in uence its sequence of synthesis. TGF-β1 is synthesized through
a series of modi cations, allowing the pre-pro-TGF-β form to evolve into a large,
latent complex (Figure 3). Starting with pre-pro-TGF-β, the signaling peptide
is removed through proteolytic cleavage, and disul de isomerase (PDI) catalyzes
the formation of three disul de bonds at Cys residues in positions 223, 225,
and 356 to allow for the dimerization of the original two monomers (Figure
Figure 3. Structural representation of the
di erent structural forms TGF-β1 takes during
synthesis, secretion, and activation.
2
e peptide
begins as a (A) pre-pro-TGF-β, transforms to a
(B) pro-TGF-β, proceeds to be a (C) SLC, later
matures to a (D) LLC and eventually is secreted as
a mature TGF-β homodimer.

33
3A). e product of this is the linked combination of two latency associated
peptide (LAP) chains and 2 mature TGF-β chains, together making up the pro-
TGF-β homodimer (Figure 3B, Figure 4).
2
is homodimer then undergoes
proteolysis catalyzed by the paired basic amino acid enzyme known as furin. is
is responsible for cleaving a bond between amino acid residues 278 and 279 which
then separates the covalent linkages holding the LAP and TGF-β chains together.
When only noncovalent forces are holding together these chains, the polypeptide
is in the small latent TGF-β complex (SLC), during which these forces maintain
TGF-β in its inactive form and prevent its interaction with receptors (Figure 3C).
Next, the SLC forms an additional disul de bond between Cys residue
33 and the Cys residue 8 of the third cysteine-rich domain of the latent TGF-β
binding protein (LTBP), leaving the complex in its large latent TGF-β1 complex
stage, or LLC (Figure 3D). e LTBP functions to form ECM proteins, and
Figure 4. Synthesis and activation of TGF-β1. 1. TGF-β synthesis begins with an inactive
precursor with a pre-signal peptide and a pro-N-terminal peptide. 2. Proteolytic cleavage
removes the signal peptide, followed by dimerization. 3. Next, cleavage by proteases
results in C-terminal mature peptides and N-terminal latency associated peptide (LAP),
forming a small latent complex. 4. e SLC transports to the extracellular matrix for
further covalent binding to latent TGF-β binding protein (LTBP), making a large latena
complex (LLC) and forms ECM proteins. 5. Activation occurs as the LLC is released from
the ECM by proteases. 6. e mature protein is then cleaved from LTBP via acidity or
plasmin, and once the TGF-β protein is released from the ECM, it can begin signaling.
34
protein activation occurs once proteases release the LLC from the ECM (Figure
4). Finally, plasmin cleaves the mature TGF-β1 protein from LTBP, freeing
the TGF-β1 homodimer and leaving it ready to begin interacting in signaling
pathways (Figure 3E, Figure 4).
TGF-β Signaling Pathway via SMAD Complexes
Once TGF-β is synthesized and activated, it may begin participating
in signaling pathways by forming SMAD complexes. SMADs are a structurally
similar protein family with the core physiological function of transducing TGF-β
signals for ultimate eects on growth and development. ese complexes then
transport to the nucleus to execute roles as transcription factors, eventually
activating TGF-β receptors to induce signaling.
5,6
Depending on where in the body
the target cells of this process are localized and which other signaling pathways
are present nearby, a variety of results can be exhibited due to SMAD complex-
based signaling, making TGF-β highly pleiotropic in nature.
7
is process begins
when TGF-β locates the surface of the cell it wishes to target and binds to the
dual-specicity kinase receptors. Structurally, these receptors highly resemble
serine/threonine and tyrosine kinases, so it logically follows that they function
through phosphorylation. Once two TGF-β molecules bind to a heterotetrameric
complex of two type II and two type I receptors (allowing for a symmetric 2:2:2
structure), the type II receptors phosphorylate the type I receptors to induce
their activation.
5,6
Depending on the specic TGF-β superfamily in action, it
will bind a specic combination of type I and type II receptors. Specic to the
TGF-β subfamily, all three members bind to the same type II receptor known
as TβRII, as well as the ubiquitously expressed type I receptor known as TβRI,
or activin receptor-like kinase 5 (ALK-5). While TβRI and TβRII are typically
found as monomers, homodimers, and heterodimers, the binding of a TGF-β
ligand preferentially induces their orientation in a heterotetrameric complex.
While TGF-β1 and TGF-β3 bind to TβRII with a stronger anity than to TβRI,
the reverse is true for TGF-β2, indicating the respective order in which each is

35
recruited for the eventual complex assembly. Speci cally, TGF-β1 associates with
TβRII to form a new high-a nity binding site for TβRI.
Next, the activated type I receptors proceed to activate the two receptor-
activated SMADs (R-SMADs), SMAD2 and SMAD3. SMAD7 functions to
compete with the R-SMADs for binding with the type I receptor, preventing
the activation and signal propagation of R-SMADs. R-SMADs and the type I
receptor then dissociate and SMAD2 and SMAD3 complex with SMAD4. is
trimeric complex of SMAD2, SMAD3, and SMAD4 all transport to the nucleus
and compound with DNA binding transcription factors (TF) and chromatin
remodeling proteins (CR), ultimately regulating target gene transcription
negatively or positively. SMAD7 has the ability to inhibit nuclear SMAD complex
transcriptional activity (Figure 5). e SMAD signaling system has key roles in
the antiproliferative properties of TGF-β as missense mutations in the pathway,
Figure 5. TGF--β/SMAD Signaling Pathway TGF-β triggers phosphorylation of
receptors upon binding, ultimately activating the R-SMADs. SMAD7 prohibits RSMADs.
SMAD7 prohibits R-SMAD activation, allowing for complexation of R-SMADs with
SMAD4. SMAD complexes ultimately associate with TFs and CRs to regulate gene
transcription.
36
prevention of phosphorylation, and blocking complex formation between
SMAD2 and SMAD3 have all demonstrated consequential tumor formation.
TGF-β1 and Cancer & erapeutic Potential for Treatment
When TGF-β is considered in the context of cancer regulation, its
paradoxical role in this process is exposed. While it stunts cancer progression
through preventing the transformation of cells in the earlier stages of tumorigenesis,
it ultimately ips its functionality and assists the processes of tumor formation
and metastasis. Specically, the protein helps promote epithelial to mesenchymal
transition (EMT), or the process by which cells lose their polarity and cell-cell
adhesion properties, allowing them to migrate and invade tissues as they transform
into mesenchymal stem cells. Additionally, TGF-β stimulates angiogenesis and
suppresses the immune system, creating a very hospitable environment for tumor
formation. TGF-β normally acts as a potent inhibitor of cell proliferation and a
tumor suppressor by prohibiting the progression of cell-cycle phase G1 through
inducing CDK inhibitor expression (p15 and p21) and suppressing c-Myc
expression. When this normal signaling pattern is disrupted, tumor formation
occurs, presenting as a variety of cancers.1 In these cells, TGF-β reverses its
functionality and works as an oncogenic factor while losing its anti-proliferative
response. Tumor cells downregulate their p15 and p21/WAF1/CIP1 signaling
pathways through interacting with the Myc/SMAD3 complex and activating the
P13K-AKT pathway.
TGF-β may act as a tumor promoter when cancer cells begin to evade the
typical inhibitory eects of the cytokine, and in turn over express its production.
is leads to uncontrolled cell proliferation and eventual tumor formation. EMT,
as described above, induces cytoskeletal rearrangements to allow for increased
cellular invasion and migration. Additionally, TGF-β is capable of increasing the
expression of vascular endothelial growth factor (VEGF) in cells, which stimulates
angiogenesis to further sustain tumor cells. Finally, TGF-β is normally responsible
for the development of a vast array of T cell lineages in the body. In highly

37
progressed malignant
tumors, it inhibits NK
cell activity, decreases
the production of
cytokines, inhibits
the development
of dendritic cells,
and in uences the
cytotoxic properties of
T cells. ese e ects
collectively promote
immunosuppression,
debilitating the body’s
ability to ght cancer
progression. TGF-
β’s tumor-promoting
ability has allowed for
its signaling pathway to be considered for a potential cancer therapy target. is
could be done in three potential ways, the rst being that TGF-β synthesis can be
directly inhibited through the application of antisense molecules. Alternatively,
TGF-β can be blocked from receptor interactions through monoclonal antibodies
and decoy receptor traps, or kinase inhibitors/aptamers which interfere with the
functioning of downstream SMAD signaling protein can be used to inhibit the
TGF-β signaling pathway (Figure 6).
Conclusions
TGF-β is a highly multifunctional cytokine that plays a vast array of roles in
the human body. It is highly integrated in a variety of signaling pathways and
works in coordination with body organs to prevent uncontrolled cell proliferation
amongst many other functions. It is now a key target for cancer prevention
Figure 6. Potential applications of TGF-β in Cancer
therapy. Several control points exist for potential
manipulation of TGF-β for cancer therapy. e methods
represented above are direct inhibition cancer therapy. e
methods represented above are direct inhibition of synthesis
through antisense molecules, blocking TGF-β from receptor
interaction with monoclonal antibodies or soluble decoy
traps, and the inhibition or the signaling pathway through
kinase inhibitors or aptamers interfering with proper function
of inhibitors or aptamers interfering with proper function of
downstream SMAD signaling proteins.
38
therapy and shows promising results in this eld. Several of these therapies are
being clinically tested today, however several challenges are also presented with
this work
10
. Specically, the duality of its pleiotropic functions and implications
in cancer ultimately results in the presentation of multiple side eects in patients
undergoing these therapies. Targeting either TGF-β proteins themselves or the
downstream product of their signaling pathways may result in the unintentional
disruption of another crucial signaling pathway in the body. In order to ll in
the gaps in this work, the intersection of TGF-β pathways and the implications
of small molecule inhibitors on these pathways must be carefully mapped and
kinetically tested. is will allow for more accurate predictions of how a particular
drug therapy may positively or adversely inuence a patient in clinical practice.
While further work must be done to verify the safety of TGF-β inhibitors in cancer
therapy, this review of the structure, synthesis, and signaling of the polypeptide
presents its growing implications in the world of cancer biology.
39
References
1. Kubiczkova, L.; Sedlarikova, L.; Hajek, R.; Sevcikova, S. TGF-β - an Excellent
Servant but a Bad Master. J. Transl. Med. 2012, 10 (1), 183.
2. Poniatowski, Ł. A.; Wojdasiewicz, P.; Gasik, R.; Szukiewicz, D. Transforming
Growth Factor Beta Family: Insight into the Role of Growth Factors in
Regulation of Fracture Healing Biology and Potential Clinical Applications.
Mediators Inamm. 2015, 2015, 137823.
3. Weiskirchen, R. BMP-7 as Antagonist of Organ Fibrosis. Front. Biosci. 2009,
14 (1), 4992.
4. Wharton, K.; Ray, R. P.; Findley, S. D.; Duncan, H. E.; Gelbart, W. M.
Molecular Lesions Associated with Alleles of Decapentaplegic Identify
Residues Necessary for TGF-Beta/BMP Cell Signaling in Drosophila
Melanogaster. Genetics 1996, 142 (2), 493–505.
5. Heldin, C.-H.; Moustakas, A. Signaling Receptors for TGF-β Family
Members. Cold Spring Harb. Perspect. Biol. 2016, 8 (8). https://doi.
org/10.1101/cshperspect.a022053.
6. Tzavlaki, K.; Moustakas, A. TGF-β Signaling. Biomolecules 2020, 10 (3), 487.
7. Derynck, R.; Budi, E. H. Specicity, Versatility, and Control of TGF-β
Family Signaling. Sci. Signal. 2019, 12 (570), eaav5183.
8. Seoane, J.; Gomis, R. R. TGF-β Family Signaling in Tumor Suppression and
Cancer Progression. Cold Spring Harb. Perspect. Biol. 2017, 9 (12). https://
doi.org/10.1101/cshperspect.a022277.
9. Haque, S.; Morris, J. C. Transforming Growth Factor-β: A erapeutic
Target for Cancer. Hum. Vaccin. Immunother. 2017, 13 (8), 1741–1750.
10. Huynh, L. K.; Hipolito, C. J.; Ten Dijke, P. A Perspective on the Development
of TGF-β Inhibitors for Cancer Treatment. Biomolecules 2019, 9 (11), 743.
40
Research Proposal: Reevaluating the Economic
Imperative to Learn
by Angelina Little
From POLSCI 381: Political Science Research Design
Nominated by Nancy Burns and Ben Goehring (GSI)
Angelina Little ask how undergraduates understand the purpose of
education, with a special interest in the role economic success plays in that
thinking. In her proposal, she works carefully and imaginatively through the
literature. She teases alternatives and distinctions from the literature that she puts
into practice in her research design. She develops a plan to interview a stratied
sample of undergraduates about these issues. She harnesses the intervention that
COVID has made in these students’ lives in her interview design. e proposal is
engaging and compelling. It outlines a research project that will be an important
contribution to the literature.
-- Nancy Burns and Ben Goehring (GSI)
Excellence in Upper-Level Writing
(Social Sciences)
41
Research Proposal:
Reevaluating the Economic Imperative to Learn
I. Introduction
What is the purpose of pursuing an education? In 375 BC, Plato wrote:
“the direction in which education starts a man will determine his future life,”
(Republic IV). Plato understood education to be a lifelong process inculcating
virtue in individuals. In our contemporary world, education is understood in the
context of the drive to succeed in a rapidly changing world. Under late capitalism,
individuals are taught to make sense of their lives in economic terms, each
decision a cost-benet analysis, each experience a potential new angle with which
to market oneself. Simultaneously, American individualism instills the belief that
each person is responsible for his own success and that anyone can make it if
he or she simply works hard enough. American individualism in the context of
capitalism thus motivates a connection between a person’s esteem and his ability
to produce material wealth. is sense of economic citizenship pervades all areas
of life, including a person’s education, valued as a path for maximizing a person’s
productivity and economic worth. As a result, students spend their high school
careers learning to measure their worth in numbers as they strive to perfect their
GPAs and SAT scores to earn admission to colleges where they will continue in a
similar fashion, crafting resumes and joining organizations that might put them
at an advantage over their peers in the race for employment. e intense pressure
to succeed academically and its implicit connection to economic success seem to
increasingly inform students’ academic endeavors, while the idea that education’s
value lies in its opportunity for individual self-actualization becomes secondary.
In my research, I hope to shed light on the extent to which the economic
rhetoric of success has trickled down into college students’ understandings of
the purposes of education. I believe that the prioritization of economic success
comes at the cost of intellectual curiosity, student wellbeing, and a true sense of
community. I hypothesize that conversations with college students will reveal such
42
values to come second to economic considerations in the way they conceptualize
the value of higher education. Additionally, the COVID-19 pandemic has
posed a unique disruption to the status quo, and has forced students, teachers,
and administrators alike to reevaluate the structure of our education system. I
believe that the dialogue on campuses regarding the purpose of education in the
wake of the pandemic will reect this, signalling an opportunity to reassess the
individualist and economic rhetoric that dominates much of the discourse on
higher education.
is research will provide unique insight into real students’ perspectives
as they strive for success in a time of crisis. Higher education faces a challenge
unlike anything it has seen before as it adapts to the COVID-19 pandemic. e
norms and values that dene institutions have been called into question as leaders
make dicult decisions regarding school reopening and confront how much is
lost when education is remote. ese challenges have led us to ask wherein lies
the true value of education and what exactly students gain in return for the hefty
tuition fees they pay. My research will inform policymakers, administrators, and
instructors at all levels by oering a window into the current climate in institutes
of higher education among students reckoning with such questions.
II. Literature Review
In 1960, eodore Schultz introduced the idea of human capital, and
with it, changed the nature of the way we view education. With the simple
statement that “skills and knowledge are a form of capital,” Schultz redened
education as an investment key to economic growth (Schultz 1961, 1). In e
Introduction of Human Capital eory into Education Policy in the United States
(2017), Laura Holden and Je Biddle document the transformation of U.S.
education policy in response to the introduction of human capital theory. Holden
and Biddle write that “educational and economic policymakers at the federal level
accepted the basic assumption of human capital theory, which was that the central
purpose of education was to increase the productivity, and thus the future earning
43
power, of the student” (Holden and Biddle 2017, 539). Under human capital
theory, education policy became a subset of economic policy, as focus turned to its
potential to maximize earnings for both the individual and the nation as a whole.
In Measuring America: How Economic Growth Came to Dene American Greatness
in the Late Twentieth Century, Andrew Yarrow identies a parallel trend in the
education sector as America came to become dened in terms of its economic
prosperity. Comparing social science and history textbooks over the course of
the twentieth century, Yarrow observes a shift away from emphasis on freedom
and equality and toward praise of America’s economic growth, with educational
materials encouraging students to “work hard to create ever more abundance
for all” (Yarrow 2010, 150). Yarrow argues that this shift moved Americans to
understand success in terms of measures of GDP, stock market, and income
rather than equity or justice. is shift had ramications for how Americans
conceptualize the world around them, even outside the realm of economics: “from
politics and social science to everyday thinking, we live in the shadow of myriad
economic metaphors. Even our personal lives are framed, seriously or tongue-in-
cheek, as cost-benet calculations” (Yarrow 2010, 196).
We can see the eects of the pervasiveness of economic terms in today’s
education system. Discussions of “investment” in education dominate much of
the discourse surrounding education policy. In Capital or people — what is the
true purpose of education?, Tanweer Ali explores the implications of human capital
theory, arguing that it “excludes a discussion of social justice and de-emphasizes
the role of education in advancing individual fullment and in strengthening
democratic values,” as well as ignoring “a view of useful human activity outside
of production within a corporate environment” (Ali 2017, 4). Ali’s insights
point to what is lost when education policy is driven by a purely economic
understanding of the education system: if education is valued for its measurable
outputs, the less quantiable benets of the education system, whether they be
individual fullment or consideration of the common good, are pushed to the
side. Education theorist Gert Biesta shares this concern throughout his writings,
44
questioning whether we have lost sight of the purpose of education altogether
as policymakers look to data drawn from easily quantiable measures (Biesta
2010). Put simply, “the danger is that we end up valuing what is measured, rather
than that we engage in measurement of what we value” (Biesta 2010). Alisdair
MacIntyre articulates a similar problem he calls the “input-output system” of
education in Alisdair MacIntyre on Education: In conversation with John Dunne
(2002). MacIntyre writes that a focus on measurable outcomes “loses sight of the
end of education, the development of its students’ powers, and substitutes for
this end that of success by the standard of some test or examination” (Dunne and
MacIntyre 2002, 4). Under the pressure to produce high-achieving individuals,
institutions become narrowly focused on test scores and other clear outputs that
can prove the success of programs. Under such models, education becomes valued
solely in terms of its quantiable outcomes. e work of Ali and Biesta shed light
on what we lose sight of when taking a strictly quantiable, measurement-based
approach to education: the values of the education system extending beyond the
production of ecient workers.
is focus on numbers and productivity in the realm of education does
not just aect policy and curricula; it trickles down to the students themselves. e
way students speak about their education reects a capital-oriented understanding
of its purposes. Agnes Callard’s essay on Liberal Education and the Possibility of
Valuational Progress (2017) compares three basic models of understanding college
education: the parental model, under which teachers take on the responsibility of
changing their students, the consumer model, which presupposes students enter
college as fully informed “consumers” seeking to maximize return on investment,
and the aspirant model, which allows for students to explore disciplines and
discover what they seek to learn with some guidance (Callard 2017). While
Callard argues in favor of the aspirant model as the best mode for students to
engage with educational materials with curiosity and openness, it seems that the
consumer model dominates most contemporary understandings of education. As
Callard points out, students pay large tuitions, “shop” for classes, and leave with
45
knowledge and abilities valued in the market; it is therefore not unreasonable for
them to view college as a “rational skills factory” (Callard 2017, 19). Inuenced
however implicitly by the advent of human capital theory amid the pressures of
an increasingly competitive market, students feel the necessity of maximizing the
return on the thousands of dollars they invest in their education. Such thinking,
while practical in today’s climate, leaves little room for the student-as-aspirant
who enters the education system as an open minded explorer and nds self-
fulllment in what he or she learns.
Biesta and Deborah Osberg make a comparable claim in Beyond
curriculum: Groundwork for a non-instrumental theory of education (2020). Biesta
and Osberg criticize contemporary understandings of education as a “curricular
instrument designed to facilitate a purpose external to itself,” advocating instead
for an emergent model that “self-generates the purpose it serves” (Biesta and
Osberg 2020, 59). Education, they argue, should not be considered an instrument
to success so much as an experience that generates meaning in and of itself. is
emergent model lines up with Collard’s aspirant model of education; neither
posits that students should use education to pursue a specic outcome, but rather,
discover that outcome as they engage in the educational process. Human capital
theory however, with its focus on education as a means of generating greater wealth,
lends itself to an instrumental model of education rather than an emergent one.
While none of these authors might put forth a comprehensive statement
on what specically education systems should aim to achieve, their analyses
demonstrate the shortcomings of today’s number-oriented approach. Whether
you call it the consumer model, the instrumental theory, or the input-output
system, the sentiment is clear: the value of one’s education is not fully realized
when academia is understood primarily as a means to an economic end. Students
and institutions alike are prone to forgoing the less quantiable benets of a more
balanced approach to education when they consider schooling to be a process
of producing capital and maximizing return on investment without regard to
personal fulllment.
46
What are the consequences of such heightened emphasis on the
economic value of one’s education? According to Michael Sandel’s account of
contemporary higher education, students lose out on more than just the ability
to explore their academic curiosities. In his book Tyranny of Merit (2020), Sandel
explores the ramications of the hyper-competitive nature of college admissions.
ough Sandel frames the issue largely in terms of meritocratic hubris, or the false
superiority students feel upon ‘earning’ admission at elite institutions, Sandel’s
account is useful in understanding the implications of measurement-focused
education models. Referencing rising cases of depression, anxiety, and substance
abuse, Sandel identies the “mental health epidemic among privileged youth” as
a consequence of students competing to achieve perfect GPAs, test scores, and
extracurriculars, explaining that “years of anxious striving leave young people
with a fragile sense of self-worth” (Sandel 2020, 181). It’s clear that even among
the most privileged students, education no longer provides a true opportunity
for cultivation of the self; amidst competition and the pressure to set themselves
apart, students strive to perfect their measurable attributes at the expense of their
own wellbeing. Sandel explains this development as “the conversion of college into
basic training for a competitive meritocracy, an education in packaging oneself
and applying for stu… the sorting and striving crowd out teaching and learning”
(Sandel 2020, 182). We can intuit that these students do not understand their
schooling as an opportunity for personal growth so much as an opportunity to
grow their human capital in preparation to enter the market. In this way we
can see some of the more dangerous consequences of teaching students to view
themselves as commodities; their self worth becomes intrinsically tied to what is
measurable and what is economically valuable.
Elsa Davidson’s interviews with teenagers at a Silicon Valley high school
in her article Marketing the Self: e Politics of Aspiration among Middle Class
Silicon Valley Youth (2008) oer further insight into the lives of students learning
to market themselves. Davidson uses interviews to examine how middle class
students are taught to understand themselves as economic citizens and how
47
“this politics of citizenship obligates middle class youth to ‘package’ or market
authentic personal traits to showcase their exceptional qualities, well-roundedness,
and authentic originality, and to frame such acts in terms of personal choice”
(Davidson 2008, 2814). In order to maximize their future earning potential,
students learn to commodify and market themselves and compete against their
peers. Davidson’s work unveils students pursuing “passions” not out of genuine
curiosity but rather as part of a performance necessary to gain access to elite
institutions and maximize their human capital. Here we see the realization of
Biesta’s theory that “the high performing school is an organisation in which the
personal is used for the sake of the functional: community is valued, but primarily
for instrumental purposes within the context of the market-place” (Biesta 2020).
Just as students perform “well roundedness” on their resumes, they might pursue
involvement in a “community” that can be packaged for a personal statement
essay. Biesta worries that if this trend continues, “it will inevitably result in the
demise of education as a holistic process of human being and becoming” (Biesta
2020). Davidson’s interviews indicate that for some students, the process of
individual exploration is restricted to what is deemed desirable on a resume. She
observes that the pressures of such performativity “suggest a domestic politics
of ‘hyper-vigilance’ that may transform young people’s self-perceptions, attitudes
towards schooling, and aspirations, while also potentially posing risks to youth”
(Davidson 2008, 2816). e lack of authenticity students experience amidst a
competitive culture with the promise of potential economic prosperity guiding
each aspirant compels students to build their self esteem from how well they are
able to “sell” themselves.
Under today’s economic impetus, students learning to market themselves
to institutions and future employers as early as high school lose out on the chance
to build meaning as they explore topics that truly interest them. Education
takes on an individualist, hypercompetitive form devoid of authenticity as
students seek only to perform rather than to truly engage in their interests or the
material. Students measure themselves in terms of the numbers that dene them,
48
understanding that this is how they will be evaluated by others. Aspiring to be the
“best” has an inexplicably material connotation as students seek to maximize their
human capital as they prepare to enter the market; under such a system, education
becomes a zero-sum game in which students compete to demonstrate marketable
skills and interests.
Such criticisms of market-oriented education beg the question of how
one might imagine a system that goes beyond measures of productivity and
excellence. In his famous address “Learning to be Human” (2012), philosopher
John MacMurray gives an idealized vision of education that extends beyond
economic success: “whilst the imperatives of earning a living… must necessarily
and properly feature in education they should not comprise the whole or even the
most important part of it. e key point is that they should be approached through
other aspects of education as part of the whole task of learning to be human in
its richest and most fullling sense” (MacMurray 2012, 661). While “learning
to be human” may at rst glance appear a luxurious goal that is not pragmatic
for educational institutions to strive for, it might hold the key for a vision of
education that values students for more than their future earning potentials.
MacMurray writes that “learning to live in a community” is the primary
purpose of education (MacMurray 2012, 662). While this may not seem radical,
when one considers this idea against the backdrop of today’s individualist,
competitive academic environment, living in a community with others does
not appear to be a primary focus. James MacAllister explains in his piece What
should educational institutions be for? (2016) that “if persons are only encouraged
to perform the practice… with a focus on their individual desires alone then it
will become all too easy for common concerns to get lost. If on the other hand
persons are encouraged to consider how their performance… might inuence the
course of their life and those of others in the longer term then it will be much
easier to raise issues concerning the public good” (MacAllister 2016, 387). A
shift of focus away from individual economic contributions and toward a public
good that is measured not just by GDP but by equity, justice, and democratic
49
values might lift reorient students to be able to learn authentically without the
pressures of marketing themselves. It would also make room for learning to
value political, social, and community-oriented contributions that cannot be
measured in terms of economic productivity. However, such a shift would require
a collective movement, as MacAllister hypothesizes that only if “parents, teachers,
school pupils, students, politicians, academics and other persons besides these do
together question the purpose of educational institutions it may become possible
to interrupt the individualistic ideals that pervade much contemporary public
life” (MacAllister 2016, 389).
e aftermath of the COVID-19 pandemic might provide a unique
opportunity to do this. e upheaval and reorganization of so many institutions
in the wake of COVID-19 has resulted in widespread critical questioning of
the purposes of the structures we have come so accustomed to. In a short piece
entitled Have we been paying attention? (2020) Biesta states that “the current crisis
has revealed with much clarity how important public education—education
funded by public means, accessible to everyone, and accountable to the public—
is, particularly for those who only have limited resources of their own. We knew
this, of course, but it is one of the more inconvenient truths about contemporary
education that we tend not talk about,” (Biesta 2020, 12). No longer are
questions about the institutional structures of education restricted to educational
philosophers and policymakers; they are instead at the forefront of conversations
between teachers, students, and parents across the country grappling with the
upheaval of the pandemic. e people are not only being reunited with the
importance of education, they are also coming to terms with uncertainty of
the future and the insecurity of jobs. Rachel Buchanan writes in Philosophy of
education in a new key (2020) that “today’s children need to be equipped with
skills rather than content in preparation for an unknown technological future”
and advocates that educators “reinvigorate education with collective thinking
and ethics” to better prepare “students for existence in the contemporary world”
(Buchanan et al. 2020). e idea that change is necessary is becoming mainstream
50
as we confront the uncertainty of the times we live in; a reframe of what is useful
when it comes to education is no longer a radical idea but entirely necessary in a
rapidly changing America.
A shift toward collective problem solving might be just what is needed
to counteract the individualist framework that has dominated the education
system in recent years. Michael Peters, Sonja Arndt and Mark Tesar claim the
pandemic has provided just the environment for such a shift by changing the
way we relate to others in their contribution to Philosophy of education in a new
key. Peters et. al write that the rules citizens now follow to limit the spread of
the virus, such as wearing masks to protect others from our own germs, are a
rst step in changing such relations: the precautions we take “indicate something
more philosophical — the ethics of the other, the ethics of care for the other, the
ethics of duty, of treatment of the other” (Peters et al. 2020). is community
orientation, under which we understand ourselves not only as individuals but as
a part of a community in which consideration of the other is essential, could lay
the groundwork for a shift away from the zero-sum, competitive understanding
of education we hold today. In e disorder of things, Peters expands upon this
idea, writing that “COVID-19 exposes the signicance of the public sector for
our collective well-being… ‘solidarity’ ‘community’ ‘collective responsibility and
action’ are the key words ringing out as a response” (Peters 2020). e shift toward
community action necessary to combatting the pandemic could be the key to a
collective overhaul of individualist attitudes driving competitive ideals of success.
e movement of rhetoric away from individuals and toward community as
witnessed by COVID-19 lays the potential for a similar rehaul of contemporary
models of education. Perhaps we could reimagine educational institutions not as
human capital producing factories, but rather, as environments for the cultivation
of the self with an orientation toward the common good. Redening success in
terms of what is good for all rather than what is good for the individual has the
potential to reunite education with its purposes beyond economic prosperity and
endow students with true fulllment.
51
e existing scholarship on the ramications of human capital theory
provides an important framework for understanding today’s education system.
Human capital theory works in conjunction with the reliance on quantiable
measures of educational success to produce a society more concerned with
measurable outcomes and wealth creation than the cultivation of the self and the
common good. I believe that students’ own understandings of the purposes of
education will reect this prioritization of economic productivity and demonstrate
the inuence of human capital theory on students’ academic decisions and
experiences. Further, I believe that the COVID-19 pandemic poses an opportunity
for a shift in the way we understand education’s purposes and value.
III. Research Design
My research will measure student perceptions of the purposes of
education and the extent to which student views are informed by individualist,
economically-oriented mindsets. I expect student perceptions to be inuenced
by a number of environmental factors such as conversations with their peers,
pressure from parents, relationships with past teachers, media consumption, and
other sources of rhetoric on education. My aim is not to pinpoint which of these
factors is most important in inuencing student views on education, but rather
to demonstrate how ideas of individualism and economic citizenship pervade the
ways students speak about why they value education. Just as under human capital
theory, education is valued as a key factor in economic productivity, I expect
students to make sense of their education as a path to success more than as an
avenue for self growth.
Because the COVID-19 pandemic has caused such a disruption to the
education system, I expect it to play a signicant role in the current discourse
on campuses regarding education. Amidst the chaos of adjusting to living in the
pandemic, many students ask what the point of paying tuition and earning a
college degree is. Some have even opted to take time o and pursue alternate forms
of education. I expect to nd that the onset of the pandemic has pushed students
52
to reconsider the purposes of education and nd the economic imperative to be
an insucient answer to the question, what is this all for?
I plan to measure students’ views on education by conducting semi-
structured interviews with undergraduate students at the University of Michigan.
I believe participant interviews provide the most eective avenue for me to fully
capture the nuanced ways in which students think about education. Each student
has a unique experience with education, and at the University of Michigan, the
nearly 30,000 undergraduates come from many dierent backgrounds. I expect
to be able to identify patterns within these conversations to get a sense of general
trends in the ways students are thinking about education in 2021. Although using
in-depth interviews will limit the number of students I am able to include in my
sample, it is essential for my research to be able to speak with students face to face
(whether that is over Zoom or in person and socially distanced) in order to get an
accurate understanding of the complex ways they conceptualize education.
e semi-structured nature of the interview will allow the students to
use their own words and speak candidly about how they think about education. I
will begin by asking open-ended questions about what they value in an education
that will allow the interviewee to lead the conversation in the direction they nd
most interesting, allowing me to take a step back and listen without leading
the conversation in any particular direction. From this I will be able to discern
patterns of what topics are at the forefront of students’ minds. I will also ask
students to describe the climates of the high schools they attended as well as why
they chose to apply to and attend the University of Michigan. eir responses to
these background questions will give me insight into the extent to which they
experience academic pressures and desire economic achievement, critical to my
analysis of their overall understanding of education’s purpose. By asking about
their previous educational environments, I will also gain understanding of the
inuence of peers and role models in their views on education, giving me a window
into how such rhetoric circulates. I will also ask how each student’s educational
values have inuenced academic decisions they’ve made, such as choice of major,

53
to discern the concrete consequences of ideological discourse. Finally, I will ask
how the pandemic might have caused them to re-evaluate any of these topics
in order to ascertain the extent to which COVID-19 may have inuenced their
views. As I collect interviews, I will review my notes and adjust my questions if I
nd recurring trends in subject matter brought up by students.
To gather subjects, I will use a snowball sampling method, reaching out
to students across disciplines who express willingness to have a conversation about
education as the ‘seeds’ of my sample and then ask them to refer me to others who
might be willing to participate in my study. I will start with several seeds that vary
in socioeconomic status, gender, race, and ethnicity in order to ensure my sample
is diverse. is sampling method will be benecial to me because it will allow me
to engage with verbose individuals willing to participate in a recorded interview,
and it will help me nd students with unique perspectives on education. e
groups I am primarily interested in speaking with are:
• First year students: e experiences of rst year students are especially
valuable to me because of their unique position as they embark on their
collegiate careers with fresh memories of their high school experiences.
First year students will have most recently completed the college
application process and will therefore likely be very in touch with their
reasoning for attending college. ey will also most easily be able to
recall and describe the environments that shaped their perceptions
on education. I hope to speak to both undeclared rst year students
and those who have a clear idea about their career ambitions. As these
students embark on their college experiences, they will have much to
consider about what they hope to get out of their higher education.
• Students in their nal year: Seniors, superseniors, and students graduating
early will be able to reect on the ways their perceptions of education
have shifted over the course of their collegiate careers and on the causes
of these shifts. ey also will be in the process of making decisions about
post-graduation plans and will be able to reect on how their academic

54
choices inuenced these decisions. I hope to speak to a range of these
students, from those with job oers to those continuing their education
and those uncertain what’s next for them. ese students will likely feel
strongly about the value of their education as they look back and prepare
to move on to the next period of their lives.
• Students taking time o: Students who choose to take time o from
school, whether to pursue alternate learning experiences, to work, to
prioritize mental health, or for any other reason are likely to have a
unique perspective on what education means to them. Students take time
o for a variety of reasons, but most are thoughtful about why taking
time away from the traditional schooling path is the correct choice for
them. During the pandemic, taking a gap year or semester seems to be an
increasingly popular choice among my peers. I’m interested to hear from
these students and compare their thoughts on the purposes of education
to those who take more traditional paths.
is sample of students will be most valuable to me because it will
represent a diverse number of perspectives from students likely to be willing
to speak about their experiences. Conducting intensive interviews with these
students will allow me to gauge the current climate of student perspectives on
the purposes of education. Because it is not logistically feasible for me to travel
to colleges across the nation to conduct these interviews, I chose to narrow my
sample to the University of Michigan. I believe that this public institution oers
a promising population of students to base my research on because of its large
number of undergraduates from across the nation and even the world. Further,
admissions standards at the university are competitive enough that I can expect to
nd students cognizant of the eects of an achievement-focused, success-driven
environment. It is important to recognize that students attending less prestigious
universities might have dierent perspectives on the necessity of earning a degree.
Even so, I think the University of Michigan is a particularly interesting institution
55
to study, given its number of dierent colleges and programs of study and its
marketing as the home of the “leaders and the best.”
One drawback of my sampling and measurement methods is that my
results will not be generalizable to the entire population of college students, and
I will not be able to verify them through external validity. However, because
the purposes of this study are to gain a window into complex and subjective
experiences, it is more important for me to be able to go in depth with some
students rather than to take a more quantitative approach that would be unable
to capture the nuance I hope to. rough my interviews, I will be able to take
into account students’ tones, ideas, attitudes, and word choice to give a nuanced
analysis of their general understandings of the value of education.
To conduct my analysis, I will transcribe recordings of my conversations
with students. I will use an a posteriori approach to develop a codebook based
around the patterns I discern across conversations. For example, I might create a
code for references to a college degree in terms of an “investment,” or a code for
expressions of changing attitudes in the context of the pandemic. After developing
this codebook, I will apply the codes to each transcript to determine the frequency
of the patterns I observe. If I am correct in my hypotheses, students’ word choice
will reveal primarily considerations of future economic endeavors and quantiable
measures of success when speaking about the academic choices they have made.
Additionally, I expect students to express uncertainty about such priorities and
possibly to even push back on their previous perceptions when speaking about
how the pandemic has aected their beliefs about the purposes of education. I will
supplement my frequency analysis with a qualitative description of the general
attitudes and tones of students interviewed.
I am condent in my hypotheses largely because of my own identity as
an undergraduate. My hypotheses in part arose from informal conversations with
my peers across disciplines expressing frustrations with the pressure to succeed
academically to the detriment of their personal wellbeing and the ludicrousness
of such pressures pervading a time of national crisis. Even if my hypotheses are
56
incorrect, however, my research will have value in providing a window into the
minds of University of Michigan undergraduates and how their perspectives have
or have not been shaped by our current circumstances. e qualitative nature of
my research will allow me to adapt my questions and focus so as to examine with
nuance the current discourse on campuses like ours.
IV. Conclusion
Students of higher education in America today are expected to learn in
hypercompetitive, individualist environments. As students work to maximize their
productivity and economic value, our education system fails to produce balanced,
intellectually curious, self-fullled students. e advent of the COVID-19
pandemic has illuminated this reality, laying the groundwork for a rehaul of the
way we think about education.
I believe that my research will make a meaningful contribution to existing
literature on the relationship between education, our political economy, and
American individualism. Understanding the way these topics intersect is essential
in considering the role of higher education in today’s world. It is important to
ask not only what we stand to gain when we tout a strong education system
as essential to our country’s success, but also what we stand to lose. It is my
hope that this research can inform education policymakers and decisionmakers in
understanding the reality of the experience of students in a candid way that course
evaluations and other structural feedback mechanisms fail to provide.
Further, I believe this research will provide insight into the impact of
COVID-19 on education. My work will illuminate how students might be
reevaluating their education in the wake of the pandemic and the dierent ways
in which this might manifest in their academic decisions. My aim is not, however,
to analyze students’ feelings about virtual learning, but rather to investigate
how the disruption of educational norms has caused students to reconsider the
value of education more generally. Does academic exploration feel like a waste of
time during a crisis or has it gained renewed importance? Have students found
57
a renewed sense of community, or do they feel more isolated and focused on
their individual eorts than ever before? Will COVID-19 change the way we
approach the classroom long after it is no longer virtual? ese questions are just
the beginning of what my research hopes to shed light on. After the pandemic
is over, its legacy will remain; my research will attempt to identify what lessons
students are taking away from it and how they are nding value in education in
this day and age.
58
V. References
Ali, Tanweer. 2017. “Capital or people – what is the true purpose of education?”
On the Horizon. 25: 4-6.
Arndt, Sonja, Tina Besley, Rachel Buchanan, Ruyu Hung, Carl Mika, Rene
Novak, Janet Orchard, Janis T. Ozolins, Marek Tesar, Christoph
Teschers, and Michael A. Peters. 2020. “Philosophy of education in a
new key.” Educational Philosophy and eory.
Biddle, Je and Laura Holden. 2017. “e Introduction of Human Capital
eory into Education Policy in the United States.” History of Political
Economy 49: 537-574.
Biesta, Gert. 2010. Good education in an age of measurement: Ethics, politics, and
democracy. London: Paradigm.
Biesta, Gert and Deborah Osberg. 2021. “Beyond curriculum: Groundwork for
a non-instrumental theory of education.” Educational Philosophy and
eory 53: 57-70.
Biesta, Gert. 2020. “Have we been paying attention? Educational anaesthetics in
a time of crises.” Educational Philosophy and eory.
Callard, Agnes. 2017. “Liberal education and the possibility of valuational
progress.” Social Philosophy and Policy 34: 1-22.
Davidson, Elsa. 2008. “Marketing the Self: e Politics of Aspiration among
Middle-Class Silicon Valley Youth. Environment and Planning: Economy
and Space 40: 2814-2830.
Dunne, Joseph and Alasdair MacIntyre. 2002. “Alasdair MacIntyre on
education: In dialogue with Joseph Dunne.” Journal of the Philosophy of
Education 36: 1-19.
59
MacAllister, James. 2016. “What should Educational Institutions be for?”
British Journal of Educational Studies 64: 375-391.
MacMurray, John. 2012. “Learning to be Human.” Oxford Review of Education
38: 661-664.
Peters, Michael A. 2020. “e disorder of things: Quarantine unemployment,
the decline of neoliberalism, and the Covid-19 lockdown crash.”
Educational Philosophy and eory.
Plato. 1943. Plato’s e Republic. New York :Books, Inc.,
Sandel, Michael. 2020. e Tyranny of Merit: What’s Become of the Common
Good? New York: Farrar, Straus, and Giroux.
Schultz, eodore. 1961. “Investment in Human Capital.” e American
Economic Review 51: 1-17.
Yarrow, Andrew. 2010. Measuring America: How Economic Growth Came to
Dene American Greatness in the Late Twentieth Century. Amherst;
Boston: University of Massachusetts Press.
60
e Social Brain Hypothesis:
An Evolutionary Explanation for Our Big Brains
by Sahil Tolia
From ANTHRBIO 368: Primate Social Behavior
Nominated by Andrew Bernard
Sahil’s work emerges as some of the most well-cited and researched
scientic writing I have encountered in a University of Michigan classroom. In this
paper, he clearly and diligently addresses the social brain hypothesis as a possible
explanation for why primates have such large brains for their body size. His paper
is even more remarkable as a self-claimed newcomer to the eld of scientic
writing; I’m not sure I believe him! I would not be surprised to encounter his
name as an author on peer-reviewed published work in the not-so-distant future.
-- Andrew Bernard
Excellence in Upper-Level Writing
(Social Sciences)
61
e Social Brain Hypothesis:
An Evolutionary Explanation for Our Big Brains
Overview
Within the animal kingdom, primates are well known for their large brains
and advanced cognitive abilities. How did these phenomena manifest over time in
the context of evolutionary pressures? In general, there are two classes of hypotheses
that seek to explain the evolutionary drivers of primate brain size. e ecological
intelligence hypothesis argues that primates evolved large brains to cope with ever-
changing environmental conditions (Dunbar and Shultz, 2017). In particular,
this hypothesis articulates that extractive foraging, or the process of locating and
obtaining hidden foods such as roots and nuts, is of utmost importance to many
primate species (King, 1986). is process requires considerable abstraction,
reection, and ne motor control, and proponents of the ecological intelligence
hypothesis posit that these cognitive demands favored primates with larger brain
sizes (Melin et al., 2014; Parker and Gibson, 1977). Similarly, the ability of early
primates to easily detect and remember the location of ripe fruit in the forest canopy
beneted from enhancing visual and cognitive skills, which in turn promoted larger
brain sizes (Milton, 1993).
In contrast, the social brain hypothesis argues that primates’ large brains
are a product of their highly complex and demanding social lives (Dunbar, 1998; S.
Roberts and A. Roberts, 2016). In recent years, further understanding of primate
social lives has provided substantial evidence in favor of the social brain hypothesis.
Here, I will review the evidence supporting the social brain hypothesis while
focusing specically on the use of cognition in competitive social contexts. I will
consider the eects of primate group size, tactical deception within groups, and
male-male competition on the evolution of larger brain sizes.
62
Background
Cognition broadly refers to the interpretation of perceived stimuli and
applying knowledge to adapt to future environmental or social scenarios (Seyfarth
and Cheney, 1990 as cited within Hopper and Brosnan, 2012). Tool use in both
the wild and captivity are well-known examples of primate cognition. For instance,
species such as bearded capuchin monkeys (Cebus libidinosus) can assess the size
and weight of rocks to use as hammers to crack open nuts (Visalberghi et al.,
2008). Similarly, C. Boesch and H. Boesch (1990) describes a higher level of
cognitive processing known as tool manufacturing among three populations of
wild chimpanzees (Pan troglodytes), where individuals would manipulate leafy twigs
and use the stems to sh for insects. Cognitive mapping skills allow primates to
discern between physical landmarks and dynamic environmental features, which is
particularly useful for species like mountain gorillas (Gorilla gorilla) to plan travel
paths to reduce energy expenditure and maximize foraging eciency (Vedder,
1984 as cited within Hopper and Brosnan, 2012).
Each of the aforementioned scenarios is highly cognitively demanding
because each requires the integration of functions from dierent regions of the
brain. For instance, tool use and manufacturing both require input from the
sensorimotor cortex (touch and feel of the tool), prefrontal cortex (overall decision-
making), and the visual cortex (Visalberghi et al., 2008). e brain is the human
body’s most metabolically expensive organ, consuming approximately 20% of our
daily energy intake (Dunbar, 1992). is suggests that natural selection in primates
has selected for brain size to increase far beyond the minimum to simply stay alive
(Dunbar, 1998).
Across the animal kingdom, brain size generally increases with increasing
body size (Seyfarth and Cheney, 2002). However, rather than measure intelligence
solely on brain size, neuroanatomists frequently refer to brain-to-body mass ratios, or
the ratio of brain mass to body mass, as a proxy for animal intelligence and cognitive
ability (Schoenemann, 2004). Primates have been found to have particularly large
brain-to-body mass ratios compared to other, less cognitively advanced mammals
63
(Jerison, 1973). Also signicant is that primates can pack in far more neurons
in their neocortex compared to other mammals. e neocortex is the part of the
brain largely responsible for controlling cognitive and social skills such as abstract
thinking and planning (Lindenfors, 2005). Although mountain gorillas have brains
that are about seven times lighter than African bush elephants (Loxodonta africana),
gorillas contain nearly twice as many neurons in their neocortices (Jabr, 2015).
Hominids like chimpanzees and humans have signicantly larger portions of their
brains dedicated to the neocortex compared to more primitive monkeys, which
results in greater overall cognitive ability (Seyfarth and Cheney, 2002).
e social brain hypothesis has been an instrumental tool in explaining how
social interactions have promoted the evolution of larger brain sizes and advanced
cognition (van Schaik et al., 2012). We can further divide social interactions into
cooperative and competitive social contexts. In group settings, primates cooperate
on a wide variety of tasks, including group hunting and territorial defense (Boesch,
1994 as cited within Hopper and Brosnan, 2012). e social brain hypothesis notes
that a large brain is needed to adequately handle the complexity of working towards
a common goal with others (Dunbar and Shultz, 2017). In contrast, competitive
social contexts are scenarios where individuals struggle over nite resources such
as food and mates. For instance, male-male competition occurs when two males
compete with each other to mate with a female (Miller and Somjee, 2017).
e social brain hypothesis, along with Byrne and Whiten’s Machiavellian
intelligence hypothesis, suggests that cognitive skills in primates were strongly
selected for due to the need for skills such as deception and exploitation within
groups (Byrne and Whiten, 1988; Hopper and Brosnan, 2012). is paper will
focus primarily on applying the social brain hypothesis to competitive social
scenarios. is will allow us to gain a better understanding of how primates
promote their individual growth, survival, and reproduction in group settings.
64
Group Size
Among primates, studies have shown that brain neocortex size is positively
correlated with group size. Dunbar (1992) compiled data on neocortex volume,
total brain volume, and mean group size for 38 genera of primates. ere was a
clear correlation between neocortex size and group size, as primate genera with
larger neocortices tended to have greater group sizes and levels of interaction
within groups. Additionally, comparisons between relatively solitary prosimian
species and “troop-making” prosimian species found signicantly larger relative
neocortex sizes among the “troop-making” species (Sawaguchi and Kudo, 1990).
To better understand the eect of group size variation on brain size within a single
primate species, Sandel et al. (2016) focused on 23 primate species where group
size changes frequently in day-to-day life. Despite a wide range of possible group
sizes, there were still strong correlations between group size and neocortex size.
Within a single primate species, groups that were larger on average also tended to
have individuals with larger neocortices.
ese ndings prompt us to think about what selection pressures led to
the evolution of large social groups and larger neocortices. e two most cited
hypotheses are that social groups allow for defense against predators and defense of
food sources against rival groups (van Schaik, 1983 as cited within Dunbar, 1992).
Each of these scenarios requires an individual to make a variety of interactions
either with conspecics or animals of a dierent species, and the individual’s
success can often be the dierence between life or death (Sandel et al., 2016).
As a result, it is likely that animals that were better suited to handle the cognitive
requirements for managing relationships within multiple groups were more likely
to succeed (Dunbar and Shultz, 2017). e larger cerebral cortex and neocortex
that followed perhaps allowed these species to engage in new social opportunities
over time, leading to more sophistication in other aspects of life (Dunbar, 1992).
65
Tactical Deception
e Machiavellian intelligence hypothesis revolves broadly around
deception and the necessity to outsmart rivals to ourish (Schmelz and Call,
2016). In Machiavellian theory, individuals develop an “awareness of awareness”,
where they realize that other members of the same species also have similar desires
(Crow, 1993). is knowledge creates inherently competitive social environments,
where individuals often resort to creative measures to outcompete others for food
or other resources (Byrne and Whiten, 1988 as cited within Dunbar and Shultz,
2017). ese measures are best dened as social manipulation techniques that
achieve small benets without putting the individual’s group membership at risk
(Humphrey, 1976 as cited within Byrne and Whiten, 1988).
Tactical deception refers to when animals try to deceive one another
by performing a normal behavior in an entirely dierent context (Byrne and
Whiten, 1988). Byrne and Whiten (1985) coined the term after observing wild
baboon (Papio ursinus) populations. e researchers noted several distinct types of
deception that allowed individuals to benet without seriously endangering their
status within the group. For instance, juvenile baboons that came across an adult
female digging for underground food began to loudly scream in a manner normally
associated with an attack from another individual. e juvenile’s mother would
confront and chase the digging female, and the juvenile would start digging and
eating the food left by the digging female. In other instances, male baboons would
seek to divert attacks from other males by intensely gazing in the distance as if a
predator were nearby (Byrne and Whiten, 1988). is would often distract the
aggressor males enough to defuse the situation. Females also sought to divert male
aggression against them by redirecting the male’s attention to an innocent third
party such as a nearby juvenile baboon (Byrne and Whiten, 1992).
Before these observations were made, tactical deception among primates
was poorly understood and thought to be only found among chimpanzees (Byrne
and Whiten, 1988). However, further observations and pooling of research showed
that tactical deception is used by nearly all families of monkeys and apes (Byrne
66
and Whiten, 1992). Wheeler (2009) presents clear evidence of tactical deception
among wild tufted capuchin monkeys (Cebus apella nigritus) using false alarm calls.
In the wild, alarm calls alert capuchins of nearby predators. However, false alarm
calls were discovered to be used primarily by subordinate monkeys to usurp control
of resources from dominant monkeys. ese calls were given more often when
food was spatially clumped and when the subordinates were well-positioned to take
advantage of startled dominant monkeys.
e Machiavellian intelligence hypothesis viewed through the lens of the
social brain hypothesis suggests that the amount of tactical deception practiced by a
primate species is directly related to the species’ brain size. is theory was validated
by Dunbar (1993), who found that, among primates, the level of sophistication in
tactical deception was signicantly related to the species’ neocortical expansion.
Further comparative studies of prosimians, New World Monkeys, Old World
Monkeys, and apes found that the extent to which species use tactical deception
is predictive of their neocortex size and overall cognitive sophistication (Byrne
and Corp, 2004). Since the neocortex controls higher cognitive functions such as
conscious thought and communication, these studies provide further evidence that
social interactions play a signicant role in primate brain enlargement and overall
cognition.
Despite the presence of tactical deception, social living is still very
common across nearly all primate taxonomic groups. is is in part because tactical
deception is often extremely subtle and carefully perfected by individuals over time
(Gavrilets and Vose, 2006). As illustrated in the previously mentioned examples,
primates usually seek to manipulate the attention of conspecics rather than overtly
attempt to control their behavior. As a result, the deceiving individuals can accrue
small benets, such as a temporary food source or lack of aggression against them,
without directly losing other benets that come with being a part of the group
(Barrett and Henzi, 2005). ese acts of deception require considerable intellect,
including the ability to understand how conspecics think and weighing the pros
and cons of social manipulation versus direct confrontation based on the scenario
67
(Byrne and Whiten, 1988). As a result, we can argue that this aspect of primate
social life has played a signicant role in increases in brain size and cognitive ability.
Sexual Selection
e social brain hypothesis can also be applied to male-male competition,
where the complex social strategies required to successfully mate has selected for
larger brains with higher cognitive abilities (Schillaci, 2006). Male-male competition
occurs frequently in multimale-multifemale primate groups, particularly through
a dominance hierarchy that allows high-ranking males to mate more successfully
than low-ranking males (Cowlishaw and Dunbar, 1991). e evolutionary pressure
of decreased access to females among subordinate males forces them to get creative.
Using data collected from multiple anthropoid species, Pawlowski et al. (1997)
found that species with relatively large neocortices more frequently employed
social strategies to subtly undermine the high-ranking male’s ability to monopolize
females. eir ndings ultimately suggested that as brain neocortex size increased,
the correlation between dominance rank and mating success decreased.
Pawlowski et al. (1997) identied three kinds of social strategies that
subordinate males could use to weaken the power dynamic in male-male
competition. Subordinate baboons can form alliances with other low-ranking
individuals to challenge a dominant male who has access to a female in estrus (Noe,
1992 as cited within Pawlowski et al., 1997). Secondly, subordinate chimpanzees
can use blu charges, where an individual charges toward another without making
contact in hopes of appearing dominant (de Waal, 1982). Lastly, baboons and
chimpanzees can form friendships with females caring for infants with the goal
of mating with the female once she reenters estrus (Smuts, 1985 as cited within
Pawlowski et al., 1997).
Each of the aforementioned strategies seeks to accomplish a similar
goal: reduce the power hierarchy within a social group that normally prevents
subordinate males from accessing females in estrus (Cowlishaw and Dunbar, 1991).
is phenomenon provides these males with a greater likelihood of reproducing
68
and passing along their genes to the next generation. e evidence that primate
species with larger neocortices had weaker correlations between rank and mating
success provides further support for the social brain hypothesis (Pawlowski et
al., 1997). In other words, the need to manage the complex dynamics found in
social interactions like male-male competition in part led to the evolution of larger
primate brain sizes.
Conclusion
On average, primates have brains that are nearly twice as large as other
mammals of similar size (Passingham, 1981 as cited within Barrett and Henzi, 2005).
Over the last four decades, research into the evolution of large brains in primates has
primarily focused on two hypotheses. e ecological intelligence hypothesis notes
that primates evolved large brains to manage dicult environmental conditions,
while the social brain hypothesis argues that primates need large brains to cope
with the computational demands of a highly complex social life (Dunbar, 1989).
e social brain hypothesis has proven to be a durable explanation for variations
in primate brain size and cognitive ability, particularly with regards to competitive
social environments. Primates with larger average group sizes tend to have larger
brain neocortices, suggesting that these animals require more cognitive processing
power when closely interacting with more individuals. In social groups with scarce
resources, numerous primate species have turned to tactical deception techniques to
“out-wit” other group members. Species that practice tactical deception have been
found to have larger neocortices and comparatively higher cognitive sophistication.
Additionally, primate species with larger brains are more likely to practice subtle
techniques to erode power hierarchies in male-male competition.
Although the social brain hypothesis has considerable support across the
eld of primatology, we must not disregard other evolutionary explanations such
as the ecological intelligence hypothesis. Studies assessing the impact of ecological
factors such as diet quality and extractive foraging on primate brain sizes have
found evidence in support of the ecological intelligence hypothesis (DeCasien
69
et al., 2017; King, 1986; Milton 1993). Rather than view social and ecological
explanations as alternatives, it is far more worthwhile to acknowledge that each
has played a signicant role in the evolution of primate cognition (Dunbar, 1992).
Further research into certain aspects of primate social life will be intriguing
for the future of the social brain hypothesis. In particular, it is well known that male
primates tend to follow female groups, which means that male and female group
sizes evolve based on dierent demands (Dunbar, 1992). While previous studies
have focused on the relationship between total group size and brain size, it would
perhaps be more reasonable to analyze female and male groups separately in the
future (Lindenfors, 2005). Lastly, much of the social brain hypothesis focuses on
the eects of social cognition on the sizes of large brain structures. However, a more
advanced understanding of microneurobiology, such as identifying specic genes
that cause brain growth upon activation, may provide new insight into how social
situations aect overall brain functioning (Dunbar and Shultz, 2007).
70
References
Barrett, L., & Henzi, P. (2005). e social nature of primate cognition. Proc. R.
Soc. B., 272, 1865-1875.
Boesch, C. (1994). Cooperative hunting in wild chimpanzees. Animal Behavior,
48, 653-667.
Boesch, C., & Boesch, H. (1990). Tool Use and Tool Making in Wild
Chimpanzees. Folia Primatologica, 54, 86-99.
Byrne, R.W., & Corp, N. (2004). Neocortex size predicts deception rate in
primates. Proc. Biol. Sci., 27 1(1549), 1693-1699.
Byrne, R.W., & Whiten, A. (1985). Tactical deception of familiar individuals in
baboons (Papio ursinus). Animal Behaviour, 33(2), 669-673.
Byrne, R.W., & Whiten, A. (1988). e Machiavellian intelligence hypotheses:
Editorial. In R.W. Byrne & A. Whiten (Eds.), Machiavellian intelligence:
Social expertise and the evolution of intellect in monkeys, apes, and
humans (p. 1-9). Clarendon Press/Oxford University Press.
Byrne, R.W., & Whiten, A. (1989). Machiavellian intelligence: social expertise
and the evolution of intellect in monkeys, apes, and humans. Oxford,
UK: Oxford University Press.
Byrne, R.W., & Whiten, A. (1992). Cognitive evolution in primates: evidence
from tactical deception. Man, 27(3), 609-627.
Cowlishaw, G., & Dunbar, R.I.M. (1991). Dominance rank and mating success
in male primates. Anim. Behav, 41, 1045-1056.
Crow, T.J. (1993). Sexual selection, Machiavellian intelligence, and the origins of
psychosis. e Lancet, 342(8871), 594-598.
DeCasien, A.R., Williams, S.A., & Higham, J.P. (2017). Primate brain size is
predicted by diet but not sociality. Nat Ecol Evol, 1, 0112.
71
Dunbar, R.I.M. (1992). Neocortex size as a constraint on group size in primates.
Journal of Human Evolution, 20, 469-493.
Dunbar, R.I.M. (1993). Coevolution of neocortical size, group size and language
in humans. Behavioral and Brain Sciences, 16(4), 681-735.
Dunbar, R.I.M. (1998). e Social Brain Hypothesis. Evolutionary
Anthropology, 6(5), 178-190.
Dunbar, R.I.M., & Shultz, S. (2007). Evolution in the Social Brain. Science,
317(5843), 1344-1347.
Dunbar, R.I.M., & Shultz, S. (2017). Why are there so many explanations for
primate brain evolution? Philos. Trans. R. Soc. Lond. B. Biol Sci, 372
(1727), 20160244.
Gavrilets, S., & Vose, A. (2006). e dynamics of Machiavellian intelligence.
Proceedings of the National Academy of Sciences, 103(45), 16823-16828.
Hopper, L.M., & Brosnan, S.F. (2012). Primate Cognition. Nature Education
Knowledge, 5(8), 3.
Humphrey, N. (1976). e social function of intellect. In Growing points in
ethology (eds) P.P.G. Bateson & R.A. Hinde. Cambridge: Univ. Press.
Jabr, F. (2015). How Humans Evolved Supersize Brains. Quanta Magazine.
Jerison, H.J. (1973). Evolution of the Brain and Intelligence. New York: Academic
Press.
King, B.J. (1986). Extractive foraging and the evolution of primate intelligence.
Hum. Evol., 1, 361.
Lindenfors, P. (2005). Neocortex evolution in primates: the ‘social brain’ is for
females. Biology Letters, 1(4), 407-410.
Melin, A.D., Young, H.C., Mosdossy, K.N., & Fedigan, L.M. (2014).
Seasonality, extractive foraging and the evolution of primate
sensorimotor intelligence. Journal of Human Evolution, 71, 77-86.
72
Miller, C.W., & Somjee, U.S. (2017). Male-Male Competition. Oxford
Bibliographies.
Milton, K. (1993). Diet and Primate Evolution. Scientic American, 269(2),
86-93.
Noe, R. (1992). Alliance formation among male baboons: shopping for protable
partners - In: Cooperation in conict: Coalitions and alliances in animals
and humans (A. Harcourt & F. de Waal, eds). Oxford University Press,
Oxford, p. 285-322.
Parker, S.T., & Gibson, K.R. (1977). Object manipulation, tool use and
sensorimotor intelligence as feeding adaptations in cebus monkeys and
great apes. Journal of Human Evolution, 6(7), 623-641.
Passingham, R.E. (1981). Primate specializations in brain and intelligence. Symp.
Zool. Soc., 46, 361-388.
Pawlowski, B., Lowen, C.B., & Dunbar, R.I.M. (1997). Neocortex size, social
skills and mating success in primates. Behaviour, 135(3), 357–368.
Roberts, S.G.B., & Roberts, A.I. (2016). Social Brain Hypothesis: Vocal and
Gesture Networks of Wild Chimpanzees. Front. Psychol., 7, 1-17.
Sandel, A.A., Miller, J.A., Mitani, J.C., Nunn, C.L., Patterson, S.K., &
Garamszegi, L.Z. (2016). Assessing sources of error in comparative
analyses of primate behavior: Intraspecic variation in group size and the
social brain hypothesis. Journal of Human Evolution, 94, 126-133.
Sawaguchi, T., & Kudo, H. (1990). Neocortical Development and Social
Structure in Primates. Primates, 31(2), 283-289.
van Schaik, C.P. (1983). Why are diurnal primates living in groups? Behavior, 87,
120-144.
73
van Schaik, C.P., Isler, K., & Burkart, J.M. (2012). Explaining brain size
variation: from social to cultural brain. Trends in Cognitive Sciences,
16(5), 277-284.
Schillaci, M.A. (2006). Sexual Selection and the Evolution of Brain Size in
Primates. PLoS ONE, 1(1), e62.
Schmelz, M., & Call, J. (2016). e psychology of primate cooperation and
competition: a call for realigning research agendas. Philos. Trans. R. Soc.
Lond. B Biol Sci., 371(1686), 20150067.
Schoenemann, P.T. (2004). Brain Size Scaling and Body Composition in
Mammals. Brain Behav. Evol., 63, 47-60.
Seyfarth, R.M., & Cheney, D.L. (1990). How Monkeys See the World: Inside
the Mind of Another Species. Chicago, IL: University of Chicago Press.
Seyfarth, R.M., & Cheney, D.L. (2002). What are big brains for? Proceedings of
the National Academy of Sciences, 99(7), 4141-4142.
Smuts, B.B. (1985). Sex and friendships in baboons. - Aldine, New York.
Vedder, A. L. (1984). Movement patterns of a group of free-ranging mountain
gorillas (Gorilla gorillaberingei) and their relation to food availability.
American Journal of Primatology, 7, 73-88.
Visalberghi, E., Sabbatini, G., Spagnoletti, N., Andrade, F.R.D., Ottoni, E., Izar,
P., & Fragaszy, D. (2008). Physical properties of palm fruits processed
with tools by wild bearded capuchins (Cebus libidinosus). American
Journal of Primatology, 70(9), 884-891.
de Waal, F. (1982). Chimpanzee politics. - Unwin, London.
Wheeler, B.C. (2009). Monkeys crying wolf? Tufted capuchin monkeys use anti-
predator calls to usurp resources from conspecics. Proc. R. Soc. B,
276(1669), 3013–3018.
74
A Review of PLA’s Sustainability as the Future of
Bioplastics
by Leah Marks
From WRITING 400: Advanced Rhetoric and Research, Writing and
Research in the Sciences
Nominated by Jimmy Brancho
Leah’s literature review invites us into the labyrinth that is plastic
recycling and degradation. Here she puts biodegradable plastics under the
microscope and improves on the existing literature with a holistic investigation
of their environmental merit. Leah shows very strong research acumen here,
making procient use of new literature and her background in materials science.
In composing this piece, she pushes her writing craft to new heights. is review
lives and breathes. e text makes inanimate plastics and invisible chemical
processes into active agents through smart choices of subject and verb. e result
is an eortless read, a sobering investigation into a cutting-edge topic becoming
more important every day.
-- Jimmy Brancho
Excellence in Upper-Level Writing
(Humanities)
75
A Review of PLA’s Sustainability as the Future of
Bioplastics
Abstract
e need to address our abounding global stockpile of plastic waste is
critical for preventing catastrophic pollution beyond repair. Two overlapping
complications that have precipitated from this crisis are the waning supply of
petroleum feedstock and the invasion of ecosystems with post-consumer plastic.
ese patterns may be attributed to the rampant production of non-biodegradable
petroleum-based polymers—materials that expend non-renewable resources and
persist in nature for decades or centuries if left unaddressed. To abate the escalation of
plastic-induced pollution, bio-based biodegradable polymers are being considered
as potential replacements for their problematic peers. At the forefront of these
eorts is polylactic acid (PLA), a starch-based polymer praised for its low carbon
footprint and similarity in behavior to commodity plastics such as polypropylene
(PP), polyethylene terephthalate (PET), and polyethylene (PE). is paper sets out
to provide an expanded investigation of the impact of increased PLA production
beyond the canonical CO2 measures on which sustainable material selection is
typically predicated. First, the sourcing of PLA feedstock is scrutinized in order
to identify its impact on food source interception and pollution from fertilizer
runo. Next, PLA’s mechanical performance and associated strengthening
mechanisms are considered in order to determine whether it is suitable for its
intended applications. Finally, PLA’s end-of-life is tracked chronologically in order
to elucidate whether/under what conditions proper degradation is able to ensue.
While PLA demonstrates promise in areas such as land eciency and mechanical
performance, there is still considerable room for improvement in its production of
toxic fertilizer runo and degradation timeline.
76
Introduction
Plastics are everywhere; in our houses, cars, and phones; in our personal
protective equipment, drugs, and implants—and in our oceans, soils, and
livestock. While plastic usage is certainly ubiquitous now, its accompanying
environmental harm cautions against a continued reliance at the same scale.
Today’s booming plastic market poses a dual threat in its rash consumption of
non-renewable resources and unbridled output of waste: 2018 alone saw 359
million metric tons of plastic waste generated, 42% of which was either littered
or sent to landlls (Silva, et al., 2020). While consistent innovation has facilitated
gradual improvement in plastic sourcing and recycling, the Covid-19 pandemic
has intercepted much of our existing progress. At this inection point of plastic
consumption, it is critical to establish plans for optimizing sustainability in order
to prevent a worst-case scenario for our planet.
Plastic can be classied in many ways, but perhaps two of the most
fundamental and potent distinctions can be found in feedstock origin and
degradability. In this case, the two opposing groups are bio-based vs. petroleum-
based, and biodegradable vs. non-biodegradable respectively. Together these
yield four distinct combinations of properties: (1.) bio-based/biodegradable,
(2.) bio-based/non-biodegradable, (3.) petroleum-based/biodegradable, and (4.)
petroleum-based/non-biodegradable. While plastics falling in the fourth group
dominate the industry today, they are accompanied by a litany of concerns such as
a large carbon footprint and inability to decompose without industrial assistance.
As landll disposal continues to abound, bio-based biodegradable plastics have
emerged as a promising alternative: a 2017 LCA conducted by Posen et al.
projected that a switch from conventional petroleum-based polymers to corn-
based biodegradable PLA would reduce greenhouse gas emissions by 25% (Posen,
2017).
Accounting for 27% of biodegradable plastic output in 2017, PLA is
considered a leading substitute for its non-biodegradable petroleum-based
counterparts such as PP, PE, and PET (Porter, 2006). Although PLA production
77
is less pollutive, this is not where the investigation should end—neither
sustainability nor material selection are based in a monolithic criterion. is
paper will therefore holistically investigate the consequences associated with
increased PLA adoption beyond the context of a conventional lifecycle analysis
(LCA). By tracing the behavior of PLA throughout its lifecycle in terms of diverse
and pertinent metrics, I plan to delineate the benets, drawbacks, and emerging
innovations that will contribute to our understanding of whether this material is
as promising a solution as its carbon footprint portends.
Feedstock and Manufacturing
PLA is a semicrystalline bio-based biodegradable thermoplastic. With
applications ranging from durable industrial machinery to sophisticated medical
implants, it may be surprising to learn that its robust lactide polymer chains are
derived from simple starches such as corn. us, to investigate PLA synthesis we
must start by assessing the risks associated with its reliance on agriculture.
Land Eciency
One pervasive concern that comes with PLA’s starch-dependence is the
depletion of our food supply: allocating food to a format not intended for eating
may seem like an inecient use of resources amid burgeoning food shortages.
However, a 2018 study conducted by European Bioplastics revealed that plants
sequestered for plastic feedstock occupied less than 0.02 percent of the global
agricultural area. is value is expected to remain constant in spite of projections
that PLA production will double between 2018 and 2023 (European Bioplastics,
2018).
Although the spatial occupation of feedstock crops is of low concern,
there is still room for improvement in how we use and treat the land on which
they are grown. To maximize land yield, researchers are investigating routes
for converting starch byproducts such as husks and shells into usable plastics.
is would enhance plastic production and quality without increasing land
dependence. One promising application of starch byproduct conversion can
78
be found in eorts to reinforce PLA with lignocellulose bers. Lignocellulose is
a biopolymer that comes from plant residues such as corn husk and sugarcane
bagasse (Tumolva, 2016). Recognized as “the most abundant renewable biomass
on earth,” these bers serve as a potential fortier of PLA (Abdel-Hamid, et al.,
2013): a 2019 study reported early success in synthesizing a PLA- coee husk
ber reinforced composite that demonstrated a 45% and 148% increase in the 59
MPa tensile strength and 3.8 GPa elastic modulus respectively from unreinforced
PLA (Collazo-Bigliardi, Ortega-Toro, and Chiralt, 2019). is discovery sets a
precedent for concurrent optimization of land yield and mechanical performance
in the eld of bioplastics.
Pollution
While a clean environmental footprint may seem inherent to plant-based
plastics, a novel 2010 case study presented the converse: in their assessment of 12
common plastics (7 petroleum-based, 4 bio-based, 1 combination), Tabone et al
uncovered PLA’s leading contribution to eutrophication. Considered a “leading
cause of impairment of freshwater and coastal ecosystems,” eutrophication
promotes the excessive growth of algae and plants (Chislock, et al., 2013). For
this to occur, one of the inputs for photosynthesis—sunlight, CO2, or fertilizer—
must be supplied in excess. An investigation conducted by Landis et al identied
fertilizer as the ingredient supplied in excess during PLA production, with high
volumes added to stimulate corn growth (Lander, Miller, and eis, 2007). When
the high phosphorous and nitrogen content in fertilizer is relayed to water systems
through runo, consequences such as algae blooms, polluted drinking water, and
hypoxia abound. Additional concern arises from the potential for algae to produce
toxic cyanobacteria responsible for killing marine life and poisoning humans.
e high rate of eutrophication attributed to PLA production
demonstrates that choosing bioplastics over petroleum-based polyolens is not
entirely favorable from an environmental standpoint. To address this concern,
scientists are looking at the possibility of incorporating recycled starch in PLA
feedstock, thus minimizing the need to produce new corn and its ensuing demand

79
for fertilizer. According to Broeren et al. (2017), using reclaimed over virgin starch
may account for up to a 60% reduction in eutrophication.
Manufactured Product
PLA is expected to be highly versatile, with demands for it to be
both exible and rigid, durable in hot and cold climates, and compatible with
injection-molding and extrusion (Masutani and Kimura, 2015). ese properties
are not necessarily inherent, but rather instilled through chemical modi cations.
Neglecting to transform the polymer in order to yield desired performance
parameters may bring about the worst case: material failure will precipitate a
ripple e ect, compromising both the PLA and any adjacent product under its
protection. Failure accounts for a super uous use of critical resources, as the
ruptured manufactured material will be prematurely sent to waste rather than
serving its intended application. us, implementing PLA with unsuitable
properties will render the energy and pollution associated with its production
grossly wasteful upon expedited disposal. We therefore must identify methods
that will fortify material properties in order to extended PLA’s lifespan.
A baseline comparison of PLA, PET, and polystyrene (PS) compiled by
Luckachan and Pillai (2011) presents PLA as comparable alternative to its non-
biodegradable, petroleum-based counterparts:
As seen in the chart, PLA is both able to withstand a larger load than both PET
and PS and demonstrate a higher sti ness. is favors the use of PLA in high
stress, load-bearing applications that desire minimized deformation. While it is
promising to see that a bio-based material is able to behave similarly—if not
Table 1. Mechanical Properties of PLA, PET, and PS (Adapted from Luckachan and Pillai, 2011).
80
better—than a less sustainable petroleum-based version in many cases, it is not
the end of the story either: PLA is not a monolith, but rather a blank canvas ready
to be modied. We should therefore aim to improve PLA on absolute terms rather
than settle for being similar to other plastics on a relative basis. With this in mind,
it is critical to aim for improvement in strength, ductility, and thermal stability so
that we are able to meet demanding design requirements.
Blending oers one strategy for modifying PLA’s mechanical properties.
In order to gure out which blends oered desired mechanical enhancements,
Narancic et al. (2018) tested ve dierent combinations of PLA blended with one
of four polymers: PHB, PCL, PHO and PBS—all of which are biodegradable and
bio-based (petroleum-based PBS was tested as well.) Each polymer was found to
bolster dierent properties at varying proportions. For instance, an 80/20 blend
of PLA-PCL induced a ~1000 MPa decrease in Young’s modulus, and an ~215%
increase in elongation. is result is indicative of the tradeos that must be
confronted when blending polymers—while a blend may enhance one property,
it can easily compromise another. us, it is evident that blends will likely serve
as one in a series of many compounding modications.
Another common route for improving the performance of PLA is seen
in the synthesis of ber-reinforced composites. Silica-based nanocomposites have
been studied for many years, with early examples reported by Paul et al in 2003.
In their research Paul was able to fabricate a novel PLA composite with layered
montmorillonite bers throughout the matrix. e addition of silicate bers
was found to enhance thermal stability and re retardancy, and serve as a more
eective gas barrier. However, it was not until the implementation of another
silicate, 2-methacryloyloxyethyl isocyanate (MOI), that researchers were also able
to claim the enhancement of mechanical properties in tandem: Chen et al (2012)
demonstrated that the addition of MOI to PLA produced a material with 4-13
times the elongation at break of pure PLA while also maintaining its high tensile
strength. While both blends and composites serve as eective enhancements
in their own right, the next stage of PLA development will involve identifying
81
synergistic combinations of these and other potential strengthening mechanisms
to create a polymer that optimizes as many properties as possible.
End of Life
Biodegradability is often perceived as a binary property—we talk
about plastics as being either biodegradable or non-biodegradable. To adopt
this perspective, however, is to ignore the nuances of polymer chains; dierent
chemical structures yield dierent properties and thus respond to designated
degradation environments dierently. We should therefore move beyond simply
asking whether a plastic can biodegrade and investigate how biodegradation varies
for dierent materials.
Biodegradation is executed in two steps: rst, polymers are broken into
smaller oligomer chains by extracellular enzymes and abiotic reactions (Luckachan
and Pillai, 2011). is process is the equivalent of cutting a pizza into slices before
eating it—the smaller the piece, the easier it is to chew and digest. Furthermore,
a shorter chain maximizes the surface area accessible to enzymes, allowing for
expedited consumption. In this latter process, enzymes convert the polymer to
either gas or mineral salts (Luckachan and Pillai, 2011). It is critical to note that
aspects of this mechanism will vary based on material properties and climate-
related variables such as pH, moisture, oxygen and light exposure, and ambient
temperature. is is where we often face a roadblock with PLA: it is falsely
assumed that just because a plastic is biodegradable it is capable of breaking down
freely and quickly in any natural setting. In reality, PLA biodegradation is heavily
reliant upon the ecosystem in which it is disposed.
Breakdown Conditions
PLA biodegradation, or “breakdown,” has been studied in three main
environments: industrial composting, water, and soil. Each environment is
compatible with PLA to varying degrees: PLA is hydrophobic and semi-crystalline,
with an average glass transition temperature (Tg) of 60˚C. With the industrial
composting route taking place in a facility, and soil and water existing in nature,
82
only the former can be expected to reach PLA’s high Tg on a consistent basis. is
means that composting provides the fastest and most direct route for melting
secondary bonds between polylactic acid polymer and pulling them chains
apart. Sangwan and Wu (2008) corroborated the time eciency of industrial
composting by showing that a 33.0*12.5*3.0 mm PLA bar could be broken down
in merely 60 days at 55˚C, as opposed to the many months typically required
for PLA to degrade on natural land. e fact that degradation in the facility was
accomplished below the Tg may be attributed to the expeditive role of ambient
moisture present during the biodegradation process.
However, time eciency is not the only criteria we should use to evaluate
the productivity and sustainability of composting. One notable downside of
industrial composting can be found in the fact that it must expend energy and
pollute to sustain its indoor climate. An LCA compiled by Rossi et al (2015)
revealed that it may cost nearly 2 kg CO2/kg PLA to transport PLA to a composting
facility and to run it. is begs the question of why we do not rely on domestic
composting as a less energy-intensive and cleaner alternative. However, a eld test
in Greece conducted by Rudnik and Briassoulis (2011) was able to subdue any
hope in this area by showing that it took at least 11 months to fully break down a
440 m PLA lm. is may be attributed to the fact that temperatures could only
reach 42˚C during the experiment, far below the Tg.
A bigger problem arises when we enlist water to break down PLA.
While few studies on this mechanism exist, those that do have reached the same
conclusion: PLA is very stubborn in an aquatic environment. For instance,
research conducted by the California Department of Resources Recycling
and Recovery (2012) revealed that PLA did not break down at all after being
submerged in the ocean for a year. is concerning result is likely attributed to
very low water temperatures and a limited population of PLA-consuming bacteria
native to aquatic climates (Haider et al, 2018). In spite of minimal success with
water degradation, biodegradation in nature is not universally ineectual; soil-
based decomposition oers a middle ground between industrial composting and
83
water-based biodegradation in its ability to be both energy and time-ecient, and
pollution-averse. In their 2011 study, Rudnik and Briassoulis (2011) reported
that PLA took 11 months to decompose in 21˚C soil. Furthermore, because soil-
based degradation takes place in nature rather than in a facility, it may be assumed
that it mandates a much smaller energy input/pollution output than industrial
composting.
Recycling Stream Contamination
While PLA is biodegradable, it can still be recycled if kept in a stream
with only pure PLA. Although this is certainly a promising way to keep PLA
out of the ocean, it does come with a notable caveat: according to Haider et al
(2018), recycled PLA is not usable in large quantities due to a change in melt
viscosity, an o-putting yellow color, and reduced lm properties. To make it to
market, recycled PLA must be combined with a minimum of 70% virgin PLA.
is diminishes the incentive to recycle PLA, as its poor properties and resulting
low demand will render it on the cheaper end of the plastic market. However, this
is not the only instance in which we encounter recycled PLA, as corrupt optical
sorting technology can yield signicant contamination in a major single-plastic
stream.
Rather than PLA being recycled with other PLA as intended, it can
often be missorted and mixed into a dierent type of recycled plastic feedstock.
For instance, a survey conducted by the European PET Bottle Platform in 2015
revealed that, on average, near-infrared (NIR) plastic sorting is conducive to a
stream contamination rate of 5-14%. is means that a stream intended to contain
only PET may include high quantities of another plastic. In the event that PLA
is wrongly directed to the PET stream, we risk compromised quality and safety
in the nal recycled product: PLA has a melting point 100˚C below that of PET,
a disparity which brings up concern for an overly-intense breakdown of PLA.
Prolonged heating of plastic well above their melting temperature will likely result
in the production of small oligomers chain through scission. Alaerts et al. (2016)
report that this will not only yield a nal product with very inconsistent viscosity
84
and processing parameters, but it will also render the feedstock an unattractive
brown color with burnt black specks. is discoloration grossly devalues PET
resin, resulting in a nal product that is likely either undesirable or unprotable.
Furthermore, Dvorak et al (2013) report that using PET feedstock tainted with
PLA in conjunction with pure PET feedstock can make processing so dicult
that it risks all plastic involved becoming unusable. us, the invasion of PLA
in recycled PET will not only create a product of suboptimal quality, but may
also instigate the unprecedented disposal of a large quantity of unused PET. e
presence of PLA in the PET stream evidently turns recycling into an act that
derails rather than promotes sustainability eorts.
To eliminate PLA from the PET stream, we must intercept PLA
contamination as early in the recycling process as possible. is is because PLA
is uniquely situated to enter the PET stream due to its inability to separate from
PET during the sink-oat stage. is mechanism is designed to isolate PET from
all other plastic in accordance with its characteristically higher density, causing it
to sink in water while the remaining low-density material oats. However, PLA
is one of the very few materials that will also sink in water and subsequently
mix in with PET. It is therefore necessary to eliminate PLA during the one step
that precedes sink-oat: NIR sorting. While Alaerts et al. propose improving the
sensitivity of NIR sortation technology used in recycling plants today, Flemish
policy agency OVAM asserts that this would result in the unnecessary disposal
of perfectly recyclable PET bottles; if a hypersensitive sorter detects even a minor
non-plastic surface contaminant such as dirt, the plastic will be ejected from the
recycling stream and sent straight to a landll. Fortunately, another option can
be found in the implementation of machine learning in optical sorting. While
this technology is very new, it has proven to avoid future sortation mistakes by
learning from past errors. Machine learning is able to consistently designate which
plastic is which, leading to a higher probability of isolating PLA to its single-
plastic stream.
ere is another route we could take in preventing the improper disposal
85
of PLA in a conventional recycling bin: correcting human behavior. A 2020 survey
conducted by Tauk et al. revealed that consumers are either not motivated or not
aware of how to properly dispose clearly labeled bio-based plastic packaging. is
is likely what is causing invasion of PLA in PET recycling bins in the rst place. To
intercept PLA contamination of PET at its origin means that we do not need to
rely on variable sortation technology to correct our mistakes. Accomplishing this,
however, is much easier said than done. While the United States Environmental
Protection Agency suggests consulting local waste management to determine how/
whether a plastic can be recycled, this method is both labor and time intensive.
Instead, studies should be conducted to understand how to teach consumers to
properly dispose of their waste. We should investigate how to optimize our plastic
labelling system to incorporate clearer and more visible disposal instructions, as
well as consider the impact of teaching this skill in schools.
Conclusion
It is undeniable that our unquenchable demand for plastic will persist for
at least the foreseeable future. With an increasing dependence on plastic, however,
comes a daunting output of waste to landlls and nature. In order to subdue
the propagation of plastic waste, we must implement technology that is able to
address it face on; it is true that recycling allocates plastic from end-of-life disposal
to a new lifecycle. However, this does not address the inevitable introduction of
plastic to nature as a symptom to improper disposal/waste sortation and rampant
littering. is is where biodegradable plastics such as PLA can prevail: with their
inherent ability to break down in the presence of bacteria, they present a promising
route to minimizing the presence of waste.
While PLA’s potential to biodegrade quickly is certainly compelling, it
is irresponsible to place all of our faith on one material property. Scrutinizing
PLA’s potential in terms of sustainability and structural integrity reveals both a
lot of promise and a lot room for improvement. While PLA does oer a lower
carbon footprint than commodity polyolens, it still struggles as one of the
86
leading contributors to eutrophication among major plastics. Furthermore, its
failure to break down in water and proclivity to contaminate the PET recycling
stream serve as major roadblocks in preventing a congregation of pollutants in our
oceans. PLA does have many redeemable qualities, including durable mechanical
properties and the ability to break down eciently in soil with and without
industrial assistance. Addressing PLA’s pitfalls while maintaining these favorable
properties will be a critical next step to take on in the coming years, with emerging
technologies such as nanober composites and precise articial intelligence-based
sortation at recycling plants leading a promising charge. No material is or will ever
be perfect. While PLA certainly poses a case for criticism, this should not outshine
its imminent promise but rather inspire continued improvement.
Works Cited
Abdel-Hamid, A., et al. Insights into Lignin Degradation and its Potential
Industrial Applications. Advances in Applied Microbiology, 82, 2018, p. 1-28.
Alaerts, L., et al. Impact of Bio-Based Plastics on Current Recycling of Plastics.
Sustainability, 10, 2018.
Bioplastics Market Data 2018. European Bioplastics Nova-Institute, 2018.
Broeren, M., et al. Environmental Impact Assessment of Six Starch Plastics
Focusing on Wastewater-Derived Starch Additives. Resources, Conservation and
Recycling, 127, 2017, p. 246-255.
Chen, B., Chien-Chang, S., and Chen, A. Ductile PLA Nanocomposites
with Improved ermal Stability. Composites Part A: Applied Science and
Manufacturing, 42, 2012, p. 2289-2295.
Chislock, M., et al. Eutrophication: Causes, Consequences, and Controls in
Aquatic Ecosystems. Nature Education Knowledge, 4, 2013.
87
Collazo-Bigliardi, S., Ortega-Toro, R., and Chiralt, A. Using Lignocellulosic
Fractions of Coee Husk to Improve Properties of Compatibilised Starch-PLA
Blend Films. Food Packaging and Shelf Life, 22, 2019.
Dvorak, R., Koisor, E., and Fletcher, J. Improving Food Grade rPET Quality for
Use in UK Packaging. WRAP UK, 2013.
Haider, T., et al. Plastics of the Future? e Impact of Biodegradable Polymers
on the Environment and on Society. Angewandte Chemie International Edition,
58, 2018.
Landis, A., Miller, S., and eis, T. Life Cycle of Corn—Soybean Agroecosystem
for Biobased Production. Environmental Science and Technology, 41, 2007, p.
1457-1464.
Luckachan, G., and Pillai, C. K. S. Biodegradable Polymers-A Review on Recent
Trends and Emerging Perspectives. Journal of Polymer Environment, 19, 2011, p.
637-676.
Masutani, K., and Kimura, Y. PLA Synthesis from the Monomer to the
Polymer. RSC Chemical Polymer Series, 12, e Royal Society of Chemistry,
2015, p. 3-36.
Narancic, T., et al. Biodegradable Plastic Blends Create New Possibilities for
End-of-Life Management of Plastics but ey are Not a Panacea for Plastic
Pollution. Environmental Science and Technology, 52, 2018, p. 10441-10452.
Narayanan, N., Roychoudhury, P., and Srivastava, A. L(+) Lactic Acid
Fermentation and Its Product Polymerization. Electronic Journal of Biotechnology,
7, 2004, p. 167-179.
PLA and PHA Biodegradation in the Marine Environment. California
Department of Resources Recycling and Recovery, 2012.
Porter, K. Ring Opening Polymerization of Lactide for the Synthesis of Poly
88
(Lactic Acid). University of Illinois, 2006.
Posen, I., et al. Greenhouse Gas Mitigation for U.S. Plastics Production: Energy
First, Feedstocks Later. Environmental Research Letters, 12, 2017.
Rossi, V., et al. Life Cycle Assessment of End-of-Life Options for Two
Biodegradable Packaging Materials: Sound Application of the European Waste
Hierarchy. Journal of Cleaner Production, 86, 2015, p. 132-145.
Rudnik, E., and Briassoulis, D. Degradation Behaviour of PLA Films and Fibres
in Soil under Mediterranean Field Conditions and Laboratory Simulations
Testing. Industrial Crops and Products, 33, 2011, p. 648-658.
Sangwang, P., and Wu, D. Y. New Insights into Polylactide Biodegradation from
Molecular Ecological Techniques. Macromolecular Bioscience, 8, 2008, p. 304-
315.
Silva, A., et al. Rethinking and optimising plastic waste management under
Covid-19 pandemic: Policy solutions based on redesign and reduction of single-
use plastics and personal protective equipment.
Singla, P., et al. Ring-Opening Polymerization of Lactide Using Microwave and
Conventional Heating. Procedia Chemistry, 4, 2012, 179-185.
Tauk, D., et al. e Paradox Between the Environmental Appeal of Bio-Based
Plastic Packaging for Consumers and their Disposal Behaviour. Science of the
Total Environment, 705, 2020.
Tumolva, T. Green Composites Using Lignocellulosic Waste and Cellulosic
Fibers from Corn Husks. MATEC Web of Conferences, 62, 2016.
89
A Handful of Walnuts
by Julia Van Goor
From ENG 325: Art of the Essay
Nominated by Jamien Delp
Julia’s meditative analysis, “A Handful of Walnuts,” is a complex, tender
and deeply honest exploration of memory. She brings both a scientic and personal
lens to her questions as she ponders her experiences with her grandmother -
an Alzheimer’s patient - her mother, and herself. Julia presses her meditation
onwards with vivid, rich scenes that are both lyrical and matter-of-fact, just as she
challenges her own memory and studies in the medical eld to determine where
(or if) a clear distinction between forgetfulness and illness might exist. Her voice
is clear, accessible, vulnerable and intentional, and her analysis as authentic as it is
surprising.
-- Jamien Delp
Excellence in Upper-Level Writing
(Humanities)
90
A Handful of Walnuts
Minuscule spines sink into my palms. I recoil at rst, but then become
impatient and tug at the oending villain. Weathered, veiny, and slightly knobby
hands gently enclose my small ngers, halting me mid-tug.
“at is not how we pick cucumbers,” my grandma admonishes. “You have to
twist and twist until it comes o naturally. Otherwise, you’ll hurt the plant.”
“But it’s pokeyyyy,” six-year-old me whines back.
“Oh, it’s not that bad. Just wipe it o - see?” And with two brisk motions
she brushes the rest of the tiny spines from the vegetable. She moves o to a dierent
patch of her vegetable garden, and I plant myself in front of the cucumber vine to start
twisting. Eventually the vine gives way, gently depositing the emerald green cucumber
into my expectant hands.
In this same patient way, my grandma taught me that while tomatoes
and corn must also be twisted, you have to pinch green beans, and you can snap
asparagus but only as long as you do it low and close to the base. I will never forget
her life lessons in vegetable picking. Partly because they are skills I’ve repeated
nearly every year in her garden and then my mom’s. But mostly because these
are my most cherished memories of my grandma. ese are also the memories I
shared while choking back tears at her funeral two years ago.
I hadn’t planned to speak at her funeral. e pastor completely blindsided
my cousins and I when he turned to us at the end of the service and asked if one of
her seven grandchildren might share a memory. We exchanged panicked glances -
not a single one of our parents had mentioned this request and we were all pretty
sure that this pastor, who none of us had met until an hour before the service, was
going o script.
Everyone, including myself, wassurprised when I, the youngest by eight
years, approached the podium. I was moved not only out of love for my grandma
and pity for my cousins, but out of frustration. So far, every memory of my
91
grandma that had been shared was about her Alzheimer’s disease. e very disease
that caused the loss of her memory, and subsequently her independence, then
her personality, and nally her life was now robbing her of her legacy. I was mad
because her nal twelve years in a memory care facility should not ablate the
woman, mother, and grandmother she was before her disease.
So, as I approached the sparse podium, I scrambled to organize my
swirling thoughts into a semi-cohesive narrative. With emotions strangling my
vocal chords, I told the story of how my grandma taught me to pick vegetables in
her garden. I concluded my impromptu speech by tearfully begging the attendees,
few of whom I actually recognized, to remember her not in a nursing home but
in her garden.
It’s possible that I was only able to remember the woman she was before
the Alzheimer’s because I hadn’t visited her at all in the past six years. It was too
painful, so I avoided it, preferring to hold on to my warm memories of summer
vegetables and games of Sequence rather than writing new memories of cold,
unmemorable walls and one-sided conversations. I told the people who actually
cared for her at the end to remember not the Gretchen Hollister they dressed and
fed every day but the Grandma Hollister who once taught me to pick vegetables.
But I am a hypocrite.
I wrote the story of my grandma’s Alzheimer’s into every single one of my
personal statements for my graduate school applications this year. I wrote about
how the devastation of a poorly understood disease inspired my future career in
biomedical research. I wrote about how the feeling of helplessness inspired me to
try to make a dierence. But I did not write about the specics. I did not write
about how her once meticulously coied hair became at and simple. I did not
write about how a warm greeting dissolved into a mere glint of recognition in
her bright blue eyes, or how even that minor acknowledgement eventually faded
away. And I certainly did not write about summers spent picking vegetables.
I chose to simplify my grandma’s legacy down to just the name of her
disease. I did the very thing I was determined to prevent when I spoke at her
92
funeral. Even though what I wrote was all true – her condition, among other
things, led me to study the molecular basis of health and disease – I still felt cheap
selling my memories for a shot at admissions.
My hypocrisy is made even worse by the fact that, based on my studies
so far, science has very unsatisfactory things to say on the topic of memory. My
molecular neurobiology course boils memory down to electrical signals, ions,
receptors, and molecules like glutamate and dopamine. I don’t understand how
miniscule and near instantaneous exchanges of molecules can become images,
words, and emotions in my mind. Words like “potentiation” and “excitatory post
synaptic potential” swirl around my head during class. I can’t make sense of how
the same ow of charges that powers my phone also determines the storage of all
of my life’s experiences.
And the cause of Alzheimer’s disease? Just accumulations of beta-amyloid
plaques and neurobrillary tangles. Small little fragments of normal proteins that
clump together inside and outside of neurons in just the right place can make
a person forget how to be a person. I can’t comprehend my memories of my
grandma’s dementia in these terms.
To make matters worse, the medical recommendations that stem from
these molecular explanations seem almost laughably feeble. I’ve watched my mom
consume walnuts by the handful after hearing a news report about their ability to
slow the progression of the disease. She’s also religiously committed to 30 minutes
of aerobic exercise a day for the same reason. at is, until the next study comes
out. Or life gets too busy. Or she just forgets.
Because that’s the thing: it’s normal to forget some things. It’s natural to
walk into your room and forget your purpose for doing so, to have no clue what
you ate for breakfast two days ago, or to blank on the word that’s on the tip of your
tongue or the name of that one relative you hardly see. But it’s not normal to forget
that your home is your home. It’s not normal to not remember which foods you
love and which you detest. It’s not normal to lose sensical language altogether. And
it’s not normal to fail to recognize your children as more than just friendly faces.
93
I can’t dene the line between occasional forgetfulness and the onset of
disease. My mom panics every time she loses her train of thought or forgets where
she left her phone. e memory of the disease that robbed my grandma of her
memory and its hereditary roots in our family haunts my mom, my sister, and I.
e onset of dementia is a distant fear of mine, but for my mom it’s an immediate
threat, lurking around the corner of every additional birthday. Even though I
reassure my mom that her forgetfulness still falls within the normal range, I don’t
know the distinction between a scattered brain and a demented brain. I don’t even
remember when my grandma’s condition crossed the line of laughable goofs, like
calling broccoli “asparagus,” to a disease that required the 24/7 care only available
in a nursing home. I wonder if I was too young to form a concrete memory of
when that imaginary line was crossed or if that line even exists at all.
My introductory psychology course provided the most compelling analysis
regarding memory formation and recall. Essentially, the more you bring a piece of
information to the forefront of your mind, the easier it will be to remember that
information later on. But here’s the catch: every time you remember that memory,
it becomes a little less accurate. at’s because with each renewed recall, you’re not
actually thinking of the original experience but instead you’re remembering the
memory based on how you most recently remembered it.
Based on that theory, it’s completely possible that the memory about
picking a cucumber I started this essay with is entirely ction by now. But is
a memory only valid if it’s true? Saying “Well, the way I remember it…” is a
nice way of covering your butt just in case the information you subsequently
provide is later found to be inaccurate. e question “do you remember…?” is
generally a much lower stakes question than “do you know…?” Our memories
aren’t expected to be perfect, but that doesn’t make them any less powerful.
ere’s something to be said for how a memory makes you feel. Your
birthday, summer vacation, home – each of these words probably triggered an
emotion as you read them. But, if I asked you to write down the exact memory that
led to your emotional connection with each word, you would probably struggle to
94
pinpoint the specics. Not because you forgot, but because the feeling triggered
by each of these words is less of an exact memory and more of an amalgamation
of past experiences unied by a shared context and overall emotional eect.
So, when I read the word “grandma,” I have the choice of remembering
the feeling of visiting her in a retirement home after she was debilitated by
dementia or the feelings associated with summer evenings in her carefully tended
garden. Neither memory is completely accurate, so in a roundabout way they
are both equally right. I know which one I want to hold on to, even if the other
remains an unwanted squatter in my fears.
e idea that we can choose which of our memories to hold on to is
unexpectedly empowering. Our brains naturally lter out irrelevant information,
choosing not to waste precious storage space on the specics of the mundane.
By making conscious decisions of what to ignore and what to remember, we can
shape which memories remain relevant. Maybe there’s some truth to the adage
“out of mind, out of matter.” e thought of your old romantic ex probably
doesn’t trigger the same tears that it may have right after the breakup. At some
point you probably stopped seeing reminders of them, stopped pulling up their
memory, and thus actively made them less relevant to your current life.
In this way we have the potential to re-write our personal histories. After
seeing my grandma’s loss of memory result in her loss of self, I began to realize
how inextricable our past is from our present. Imagine the power that purposefully
shaping your memories could give you. Instead of remembering my failures, my
awkward blubbering, and my disappointments, I could prioritize the moments of
success, grace, and joy. Maybe instead of fearing forgetfulness I could harness its
power to build a happier history and more condent present.
I don’t have control over every experience in my life. And no matter how
many walnuts I eat, I also don’t control (or even fully understand) the molecular
mechanisms that determine the functions of my brain. But I can control the
power that my memories hold over me. I can choose to remember the grandma
who taught me to pick vegetables.

